![]()
Азендохзавр (лат. Azendohsaurus) — примитивный архозавр позднего триасового периода из Марокко и Мадагаскара. Известен лишь по зубам и челюсти. Ранее считался родом семейства фаброзаврид или прозауропод. Считается трилофозавром с 2010 года.
Описание
{kind=link}
Azendohsaurus раньше классифицировался как растительноядный динозавр, сначала как орнитихия, но чаще как прозауропод. Это было основано только на его челюстях и зубах, которые имеют общие черты, характерные для травоядных динозавров. Полный скелетный материал с Мадагаскара, однако, обнаружил более характерные черты, наследственные от Archosauromorpha, и что Azendohsaurus вовсе не был динозавром. Вместо этого Azendohsaurus был фактически более примитивным архозавроморфом, который конвергентно развил многие черты челюстей и скелета, характерные для более поздних гигантских динозавров-зауроподов. Было обнаружено, что он является членом недавно признанной группы специализированных, в основном растительноядных, архозавроморфов, которая получила название Аллокотозаврия. Это также тезка и характерный признак его собственной семьи аллокотозавров, Azendohsauridae; первоначально единственный член, семья теперь включает других подобных аллокотозавров, таких как более крупный рогатый азендозаврид - шрингазавр из Индии.

{kind=link}
Несколько других групп архозавроморф также также породили травоядных животных в триасе, иногда с подобными динозавру зубами, что также вызвало путаницу в их классификации. Azendohsaurus известен, однако, также сходящимся развитием формы тела, подобной зауроподоморфам в дополнение к его челюстям и зубам. Азендохзавр и зауроподоморфы, вероятно, независимо развились, чтобы заполнить подобную экологическую нишу как длинношеих, относительно высоко растущих травоядных животных в их среде. Тем не менее, азендохзавр предшествует крупным позднеприасовым зауроподоморфам, на которые он похож на несколько миллионов лет, и не развивал подобные планы тел в тех же условиях окружающей среды. Тогда, возможно, он был одним из первых травоядных животных, которые сыграли важную роль, которую, как считалось, занимали только крупные зауроподоморфы во время триаса, расширив известное экологическое разнообразие травоядных архозавроморфов за пределами динозавров в триасовом периоде. Azendohsaurus также важен, поскольку он может быть одним из самых ранних известных эндотермических архозавроморфов, и предполагает, что теплокровный метаболизм был наследственным для более поздних архозавров, включая динозавров.
{kind=link}
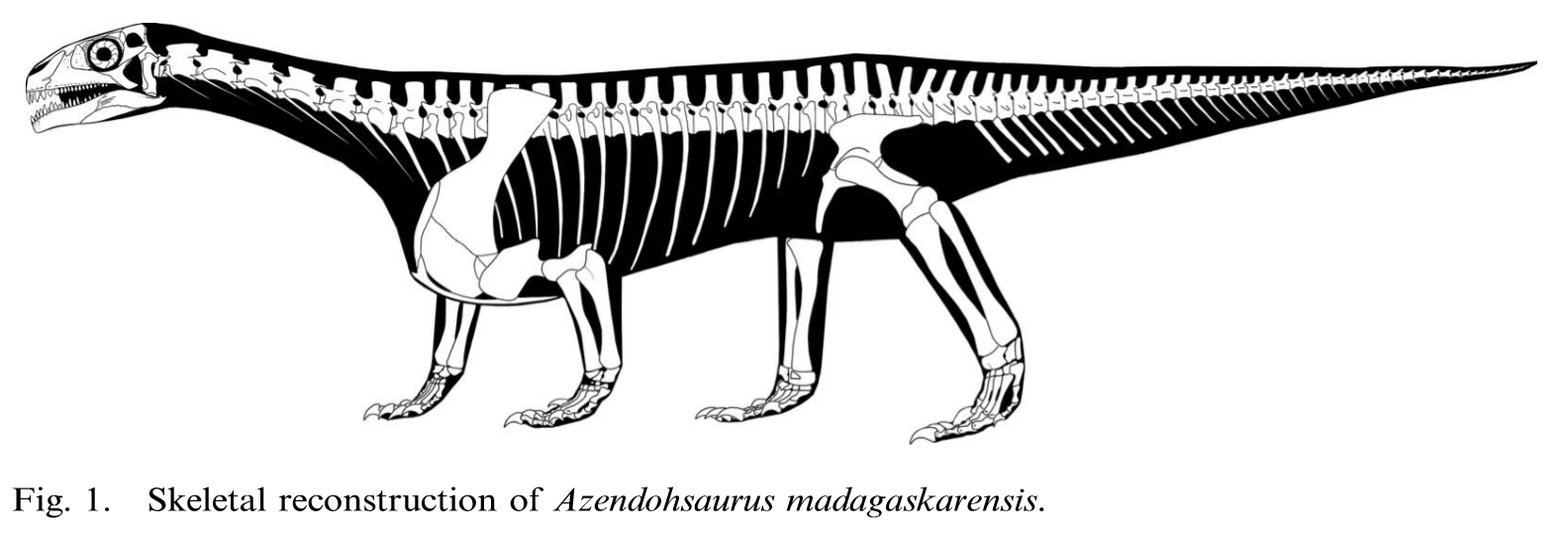
Азендохзавр был коренастой рептилией среднего размера, его длина составляла примерно 2–3 метра (6,6–9,8 фута). У него была маленькая коробчатая голова с короткой мордой на длинной шее, которая была поднята над плечами. Тело было широким, с бочкообразной грудью и плечами, намного выше бедер, вместе с необычно коротким хвостом. Его поза была полурастянутой, с раскидистыми задними конечностями и слегка приподнятыми передними конечностями. Сами конечности относительно короткие и особенно крепкие, с цифрами, которые короче и толще по сравнению с другими ранними архозавроморфами, у каждого из которых заметно большие изогнутые когти на всех четырех ногах. Внешне его внешность сравнима с таковой у зауроподоморфных динозавров, наряду с различными деталями его скелета, что позволяет предположить, что азендозавры сходятся по сходным признакам для относительно интенсивного растительного и травоядного образа жизни. A. laaroussii плохо известен по сравнению с A. madagaskarensis, и известно, что эти два вида отличаются лишь незначительными деталями челюстных костей и зубов. Дополнительный скелетный материал A. laaroussii был получен из типового местонахождения оригинальных фрагментов черепа, но до 2015 года еще не был официально описан.
Череп

{kind=link}

{kind=link}
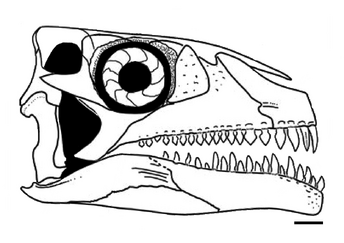
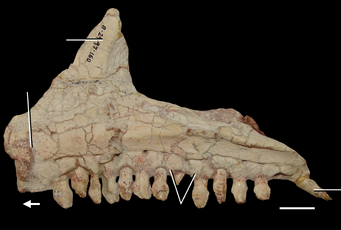
Череп A. madagaskarensis почти полностью известен, он крепко сложен, имеет короткую квадратную форму и глубокую морду. Предчелюстные кости мягко изогнуты в передней части верхней челюсти, образуя тупой, круглый кончик рыла, в то время как нижние челюсти имеют глубокий, наклоненный вниз наконечник, как у зауропод. Костные ноздри слиты в едином (сливном) отверстии, которое направлено вперед в передней части рыла, как у носорогов. У черепа есть ряд черт, придающих азендохзавру свойство с зауроподоморфами, включая изогнутый вниз зубной кость, крепкий дорсальный отросток верхней челюсти и несколько особенностей зубов. Процесс на верхней челюсти обычно указывает на наличие анторбитального фенестра у архозавров, но у Azendohsaurus это пространство занято слезной костью перед глазами. Это уникальное расположение неизвестно у других триасовых архозавроморфов, за исключением родственного Shringasaurus. Орбиты почти полностью заняты большими склеротическими кольцами, наводящими на мысль о больших глазах. Нижний височный фенестр открыт снизу, отделяя кости югала и квадратажуги (примитивная черта для архозавроморфов). Также, как и у других ранних архозавроморфов, у Azendohsaurus имеется небольшое (3–5 мм в поперечнике) теменное отверстие («третий глаз») на крышке черепа.
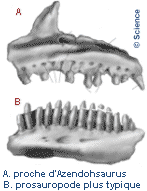
Нижняя челюсть имеет много сходств с таковой у зауроподоморфов с суставом, в котором шарниры челюсти расположены ниже уровня зубного ряда и ниспадающих зубных костей, а также зубов аналогичной формы. Эти особенности по-разному встречаются у других растительноядных триасовых архозавроморфов, но эта комбинация известна только у Azendohsaurus и sauropodomorphs. Зубы все приблизительно в форме листа (ланкоелат) с расширенными коронками и выпуклыми основаниями, которые срослись с костями челюсти (ankylothecodont). Тем не менее, верхние и нижние зубы отчетливо разнородны, и их легко отличить друг от друга. Верхние зубы относительно короткие и широкие в основании, с 4–6 зубчиками на каждой поверхности, похожие на орнитицианы; нижние зубы почти в два раза выше и имеют вдвое больше зубчиков, более близко напоминающих зубы зауроподоморфов. Четыре предчелюстных зуба являются самыми длинными зубами в верхней челюсти и имеют более изогнутую форму, чем остальные.
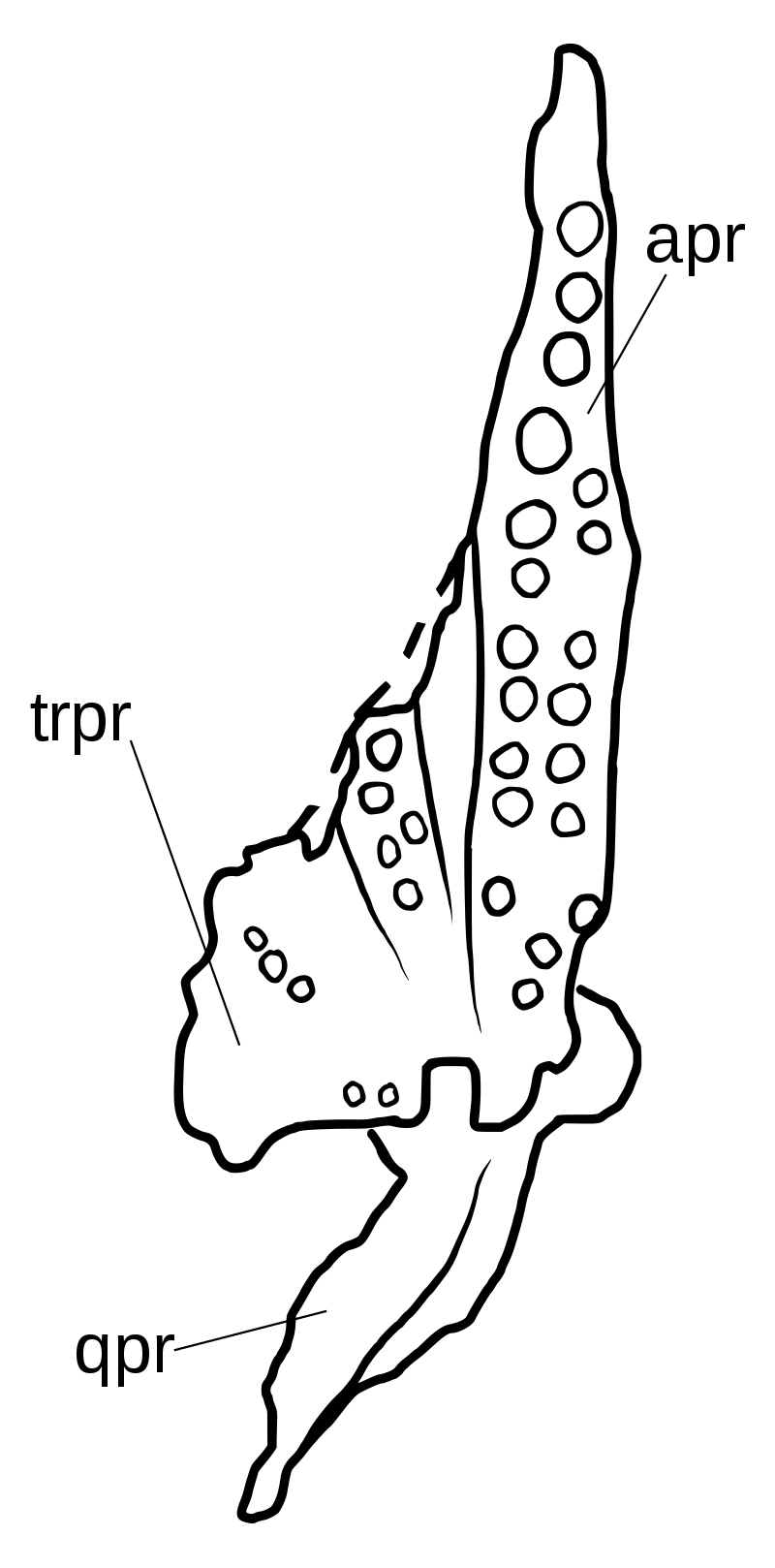
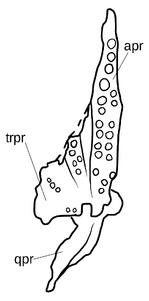
{kind=link}
Небо покрыто многочисленными полностью развитыми небными зубами, с четырьмя наборами на крыловидной кости и дополнительными рядами на небе и сошниках. Зрелые Azendohsaurus madagaskarensis имели не менее 44 пар небных зубов, в дополнение к 4 зубам в каждой челюсти и по 11–13 в верхней челюсти, а также максимум 17 зубов в зубной полости. Небные зубы нередки у травоядных рептилий, но у азендохзавра они почти идентичны по форме с краями челюсти, но немного более толстые. Другие архозавроморфы с небными зубами имеют гораздо более простой небный зубной ряд с маленькими куполообразными зубами. Тератерпетон - единственный другой архозавроморф с такими же хорошо развитыми небными зубами. Единственным описанным материалом A. laaroussii являются зубные, верхнечелюстные, предчелюстные и несколько зубов. Они широко напоминают A. madagaskarensis в общем виде, но с несколькими отличительными отличиями. Количество зубов A. laaroussii выше, с 15–16 зубами в верхней челюсти по сравнению с 11–13 зубами A. madagaskarensis. Зубы A. laaroussii также более высокие, чем у A. madagaskarensis, и имеют более плотные зубцы. Еще одним отличием этих двух видов является наличие выступающего киля на внутренней поверхности верхней челюсти. Этот киль проходит по всей длине верхней челюсти у A. laaroussii, но встречается только вдоль задней его половины у A. madagaskarensis. Любые другие возможные различия между двумя видами не могут быть определены без остальной части черепа и скелета.
Скелет

{kind=link}
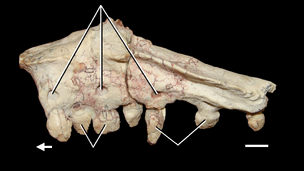
Вся известная информация о посткраниальном скелете азендохзавра основана на данных A. madagaskarensis. У азендохзавра известна большая часть позвоночного столба и хотя она неполная, по оценкам, она имеет 24 пресакральных позвонка (включая атлас и ось). Крестец бедер имеет только два позвонка, а полное число хвостовых позвонков в хвосте неизвестно, но, по оценкам, оно составляет всего около 45–55 (низкое значение для архозавра). Шейные позвонки изменяют форму вниз по шее, начиная с характерного удлинения с длинными и низкими нервными шипами, и постепенно становясь короче по направлению к основанию шеи, но с более высокими и более узкими нервными шипами. Это укорочение наблюдается у других аллокотозавров, таких как трилофозавр, но не наблюдается у других длинношеих архозавроморфов. По-видимому, азендохзавр держал шею поднятой над телом, о чем свидетельствует наклонный угол зигапофизов, соединяющих каждый позвонок, а также то, что передние зигапофизы каждого позвонка выше, чем задние. Шею также, вероятно, держали в легкой дуге, основываясь на сочлененном наборе шейных позвонков в этом положении.
Спинные позвонки обычно напоминают последние шейные позвонки с высокими вертикальными нервными шипами. Эти позвонки также уменьшаются в длину вниз по спине, но менее резко, чем в области шеи. Последний спинной, однако, уникален, так как имеет нервный отдел позвоночника, наклоненный вперед. Из двух крестцов первый позвонок является более крупным и более крепким, с высокими нервными шипами над его задней половиной. Оба крестца имеют большие ребра, полностью сросшиеся с позвонками, которые сочленяются с подвздошной костью. Хвостовые позвонки напоминают другие позвонки, но с отогнутыми назад нервными шипами. Длина хвостовых плавников и высота нервных отростков постепенно уменьшаются в хвосте, в отличие от некоторых других архозавроморфов, у которых позвонки вытягиваются к кончику. Это означает, что хвост был коротким и не сужался, но сам кончик хвоста неизвестен. У них последовательно есть шевронные грани от 3-го или 4-го позвонков до последнего каудала. Шейные ребра длинные и тонкие, становятся более крепкими и сужающимися при движении вниз по шее. Некоторые из шейных ребер из нижней половины шеи имеют небольшую грань на внутренней поверхности на их кончиках, которые могли удерживать кончик предыдущего ребра, образуя серию жестких шейных ребер (также возможно для длинного шеи танистрофея), который мог бы укрепить шею. Ребра туловища длинные и изогнутые наружу, что указывает на то, что у Азендохауруса был широкий и глубокий бочкообразный сундук. Длина и кривизна ребер уменьшается вниз по позвоночнику, а последнее ребро короткое, полностью срослось с последним спинным позвонком и указывает прямо наружу в стороны.
Конечности
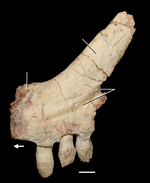
{kind=link}
Передние конечности и плечи (грудной пояс) азендохзавра хорошо развиты и крепки. Лопатка (лопатка) длинна, примерно в два раза выше ее ширины, соответствует длине и кривизне ребер, чтобы вместить глубокую грудь. Лезвие вогнутое с каждой стороны со слегка расширенным кончиком, который заострен назад. Межключичная клетка большая и крепкая и разделяет с трилофозаврами и некоторыми ринхозаврами длинный «подобный веслу» задний отросток, который сплющен и расширен к кончику. Он также обладает уникальным направленным вперед процессом, функцией, которой он обладает только у Protorosaurus и некоторых ранних диапсид.
Коракоиды крупные и округлые, сочленяются с лопаткой, образуя гленоид (лопатка). Гленоид направлен вбок, что типично для рептилий с широким телом, однако лопаточная часть направлена немного назад, что может указывать на то, что плечевая кость удерживалась в более приподнятом положении. Сама плечевая кость большая и широко расширена на обоих концах, оставляя относительно узкую «талию» в середине стержня с очень хорошо развитым дельтопоральным гребнем. Радиус также коренастый с немного расширенными концами, в то время как локтевая кость значительно расширена с обоих концов, хотя в меньшей степени дистально.
Бедра (тазовый пояс) не такие длинные, как плечевые кости, при этом три бедра примерно равны по размеру. Иллиум высокий и изогнутый вдоль верхней поверхности, с коротким округлым отростком спереди и более длинным сужающимся за ним процессом. Лобок направлен вниз и немного вперед, и имеет лишь слегка утолщенное расширение (ботинок) на конце. Считается, что седалищная кость относительно короткая, короче илий и имеет приблизительно треугольную форму с прямыми краями и закругленным задним кончиком. Поверхности сочленения между каждой седалищной костью необычно расширены по сравнению с другими архозавроморфами. Все три способствуют формированию глубокого округлого вертлужного впадины (тазобедренного сустава). В отличие от открытого гнезда динозавров, внутренняя стенка вертлужной впадины у Azendohsaurus является твердой костью.
Большие сакральные ребра сочленяются с подвздошной костью, так что она удерживается почти вертикально, с небольшим наклоном вниз, при этом тазобедренный сустав направлен наружу от тела. Бедро длинное и неопределенно S-образное, со слегка расширенной головой, которая не повернута внутрь, в отличие от динозавров, что указывает на то, что она не удерживалась вертикально. Бедренная кость также скручена вдоль ее стержня, так что грани головы и колена смещены относительно друг друга примерно на ~ 75 °. Длина бедренной кости примерно 75%, слегка изогнута и очень прочна по сравнению с другими архозавроморфами, за исключением самых крупных ринхозавров. Малоберцовая кость, напротив, тонкая и более заметно изогнута по всей длине.
Конечности Azendohsaurus хорошо представлены в окаменелостях, включая полную руку и ногу. Все запястья и лапчатые кости хорошо окостеневшие и отчетливые, а сложный лапок состоит из девяти костей. Пястные кости в руке заметны, поскольку они расходятся по гладкой дуге, причем длина цифр почти симметрична вокруг длинной третьей цифры, а также относительно не расходящихся первой и пятой цифр. Это контрастирует с руками других рептилий, где первая и пятая цифры разнесены друг от друга, а четвертая цифра самая длинная. Плюсневые кости и цифры стопы также расходятся по плавной дуге, но в отличие от кисти они не симметричны, с длинным четвертым пальцем ноги и коротким пятым пальцем с крючковатым когтем.
Все цифры рук и ног необычно короткие для archosauromorph, в отличие от родственного Trilophosaurus. Когти (или негодяи) все очень большие, узкие и резко изогнуты, и значительно больше, чем предыдущая кость пальца, к которой они были прикреплены. Пальцы и когти имеют сходные черты с дромеозаврами и манирапторами, а также с другими рептилиями, такими как черепаха проганохелис. Эти общие черты связаны с хорошо развитыми сухожилиями сгибателей, и предполагается, что это приспособление для противостояния силам, связанным с копанием.
История изучения
A. laaroussii
Первые открытия

{kind=link}
Первые окаменелости Azendohsaurus laaroussii были обнаружены в северной части формации Аргана (также известной как формация Timezgadiouine) в Марокко, часть бассейна Арганы около Атласских гор. Окаменелые пласты состоят из песчаников и аргиллитов из красной глины и были раскопаны Жаном-Мишелем Дютюитом в период с 1962 по 1969 год. Окаменелости A. laaroussii известны только из одного слоя в пласте, в обнажении, которому Дуит давал номер XVI в основание T5 (или Irohalene). T5 традиционно датируется ранним позднетриасовым возрастом с использованием биостратиграфии позвоночных, основанной на присутствии фитозавра «Paleorhinus» magnoculus, как части карнского датированного зоной Palaeorhinus, хотя этот метод -корреляция и датировка глобальных триасовых последовательностей может быть неточными, и дата для T5 остается неопределенной. Первые окаменелости состояли только из частичного зубного фрагмента зубного ряда и некоторых связанных зубов. Этот материал был обнаружен Дж. М. Дютуитом в 1965 году и описан в 1972 году, который полагал, что он принадлежит к травоядному орнитисхианскому динозавру, а также к одному из старейших динозавров, открытых до сих пор. Он назвал род «Ящерица Азендох» в честь соседней деревни Азендох, расположенной всего в 1,5 км к западу от места, где были обнаружены окаменелости. Описание Dutuit's Azendohsaurus как орнитихии было скоро оспорено палеонтологом Ричардом Тулборном два года спустя в 1974, который был первым, чтобы предположить, что Azendohsaurus был "просавроподом" вместо этого. Тот же вывод был сделан Жозе Бонепартом после изучения материала в 1976 году. Эта повторная идентификация была одобрена исследователями в последующих публикациях, и она была по-разному отнесена к семейству "просавропод" Anchisauridae и Thecodontosauridae без дальнейшего объяснения. Сам Дютюит даже согласился с тем, что Азендохсаурус, вероятно, будет «прозавроподом» в 1983 году, хотя незадолго до этого в 1981 году он кратко расценил его как «предорнитискийец». В 1985 году палеонтолог Питер Гальтон предположил, что оригинальный материал Dutuit "Azandohsaurus [sic]" включал челюсть "prosauropod" и зуб fabrosaurid ornithischian (ныне несуществующей группы ранних орнитишианов), основываясь на различиях в форме зубы. Это предложение было опровергнуто Франсуа-Ксавье Гофром в 1993 году, когда он переписал материал, а также описал дополнительные кости челюсти и зубы, включая две верхние челюсти. Он правильно сделал вывод, что материал принадлежал одному таксону, но назначил род "Prosauropoda" incertae sedis, основываясь снова на характеристиках челюстей и зубов. Тем не менее, он не смог определить его положение в «Prosauropoda» из-за неоднозначного распределения этих признаков у ранних травоядных динозавров, а также из-за отсутствия каких-либо сопоставимых триасовых рептилий, поэтому он отнес это к incertae sedis. Его оценка была принята многими другими исследователями в последующие годы вплоть до описания нового материала с Мадагаскара.
Более поздние исследования
В 2002 году на ежегодной конференции Общества палеонтологии позвоночных Nour-Eddine Jalil и Fabien Knoll были представлены новые материалы из типового местонахождения A. laaroussii, включая части посткраниального скелета. Дополнительный материал включал пресакральные позвонки, кости конечностей и пояса конечностей. Материал был разобщен, и относился только к A. laaroussii из-за его связи с узнаваемыми фрагментами черепа и челюстей. Посткраниальный материал был признан нединозаврийским, но все еще считался орнитодиранским архозавром, связанным с динозаврами. Если это правильно связано с челюстями и зубами, это указывает на то, что Azendohsaurus не был тесно связан с какими-либо травоядными динозаврами, несмотря на их сходство. Точно так же зубы от других предполагаемых триасовых орнитишей позже были найдены принадлежащими ранее непризнанным травоядным рептилиям, таким как псевдосучевий Revueltosaurus, что подчеркивает возможность ошибочной идентичности у других предполагаемых травоядных триасовых динозавров, включая Azendohsaurus. Новый постчерепной материал от A. laaroussii был описан как часть докторской диссертации Халдуном в 2014 году, но по состоянию на 2019 год этот тезис еще не был опубликован, и материал официально не описан в литературе. Тем не менее, теперь он стал более уверенно относиться к A. laaroussii после описания материала на Мадагаскаре, и было обнаружено, что он имеет как минимум две диагностические посткраниальные особенности с видами мадагаскаров. Весь материал A. laaroussii, включая голотип и неопубликованные постчерепные книги, размещен в Национальном музее естественной истории в Париже, Франция.
A. madagaskarensis
Первые описания
В 1997–1999 годах окаменелости нового архозавроморфа были обнаружены международной экспедицией на юго-западе Мадагаскара и были обнаружены в течение следующего десятилетия. Окаменелости были найдены в единственном костяном русле, называемом М-28, толщиной всего в десятки сантиметров на 100-метровом участке обнажения, обнаженном в виде возвышенной террасы реки недалеко от восточного берега реки Малио, недалеко от него. Национального парка Исало, к северо-западу от города Ранохира и к востоку от Сахахары. Населенный пункт находился у основания средне-позднейшей триасской свиты Макай, также называемой Исало II, частью группы Исало в бассейне Морондавы.
Ранее считалось, что формация относится к возрасту от ранней до средней юры, хотя более раннее открытие ринхозавра Исалорхинхуса пересмотрело эту оценку до среднего триаса. Окаменелости тетрапод, найденные в экспедиции 1997–1999 гг., Подтвердили, что Исало II является триасовым по возрасту, но вместо этого предложили более молодой карнский возраст. Возраст пласта также был соотнесен с зоной сборки Santacruzodon (AZ) из свиты Santa Maria в Южной Америке на основе общих родов traversodontid cynodonts, с аналогичным возрастом позднего ладинца или раннего карна. Santacruzodon AZ был более надежно датирован с помощью радиоизотопного датирования U-Pb, что предполагает максимальный возраст отложений 237 ± 1,5 миллиона лет в раннем карне.
Костяное ложе содержало материал почти от «десятка» особей разных возрастов и размеров, все от одного вида. Материал был очень хорошо сохранен, обычно сохраняя трехмерную форму костей с очень небольшим раздавливанием или искажением в некоторых образцах. Основываясь на состоянии сохранности, считалось, что некоторые кости были быстро захоронены, в то время как другие дольше оставались на поверхности, где их выветривали, трескали и, возможно, растоптали перед захоронением.
Как и в случае A. laaroussii, зубы и челюсти были первым материалом, который был извлечен и описан из костного ложа. Первоначально они были ошибочно причислены к двум разным видам, основываясь на разнице в форме зуба в верхней и нижней челюстях, но один из них был признан очень похожим на A. laaroussii, разделяя киль на внутренней поверхности верхней челюсти и расширенный лист зубы, среди других особенностей. Как и A. laaroussii, оба этих предполагаемых вида были также ошибочно идентифицированы как «просавроподы»; вид Azendohsaurus или родственный таксон и другой, более типичный "просавропод". Дальнейшие открытия связанного материала прояснили, что весь материал челюсти и остальная часть скелетов были от единственного, нового вида Azendohsaurus.
Переосмысление


{kind=link}
Даже предварительное исследование остальной части черепа и скелета мадагасканского вида подтвердило, что Azendohsaurus не был динозавром, а вместо этого был аберрантным травоядным архозавроморфом, который был только отдаленно связан с динозаврами, не говоря уже о зауроподоморфах. Описание черепного материала было впервые опубликовано в мае 2010 года Джоном Дж. Флинном и его коллегами, которые также официально назвали и диагностировали его как новый вид Azendohsaurus, Azendohsaurus madagaskarensis, названный в честь страны его происхождения. Это также был первый раз, когда Azendohsaurus был определен как не-динозавр в опубликованной литературе.
В декабре 2015 года Стерлинг Дж. Несбитт и его коллеги официально описали и опубликовали остальную часть скелета A. madagaskarensis, предоставив первый подробный анализ полной анатомии Азендохсауруса из ныне почти полностью известного скелета. В дополнение к сравнению его анатомии, они также смогли проанализировать его эволюционные отношения с другими триасовыми рептилиями в филогенетическом контексте впервые.
Несбитт и его коллеги охарактеризовали сохранение материала как «в целом превосходное», а количество перекрывающихся материалов облегчало определение исходной морфологии по искаженным и сломанным костям. Большая часть материала была найдена вычлененной, а иногда и изолированной, но ряд специфических частей тела был найден сочлененным в жизненном положении, включая участки шеи, спины, рук и ног. Большая часть материала была одинакового размера, с диапазоном около 25% между самыми маленькими и самыми большими образцами, хотя значение этого не понято, и это может быть связано с онтогенезом, индивидуальной изменчивостью или половым диморфизмом.
Хорошо сохраняющая природа большей части материала также позволила переосмыслить части скелета других архозавроморфов, таких как трилофозавр. Руки других архозавроморфов часто плохо известны, и поэтому их сохранение у Azendohsaurus считается важным для понимания их эволюции в ранних архозавроморфах. Все образцы A. madagaskarensis постоянно хранятся как в Университете Антананариву на Мадагаскаре (включая голотип), так и в Полевом музее естественной истории в Чикаго, штат Иллинойс, включая слепки некоторых оригинальных образцов.
Классификация


{kind=link}
Тератерпетон - представитель группы трилофозавров.
Попытки идентификации
Azendohsaurus был впервые ошибочно идентифицирован как птицетазовый динозавр Dutuit, что основывалось на общих характеристиках его зубов, таких как листообразная форма и количество зубчиков. Позже другие исследователи полагали, что это зауроподоморф, вместо этого он был отнесен к несуществующему инфракрасному отряду «Prosauropoda» (тогда считавшемуся отдельной монофилетической группой, связанной с зауроподами, теперь известной как парафилетическая степень), основываясь на морфологии нижней челюсти. , верхняя челюсть и зубы, такие как нисходящий изгиб зуба и отсутствие предзубной кости, одна из характерных черт орнитихий. Эти ошибочные определения были вызваны сближением формы челюсти и зуба между ним и травоядными динозаврами, в то время как его истинные филогенетические связи не могли быть реализованы из-за отсутствия других костей черепа и скелета.
На не-динозавровую идентичность Azendohsaurus впервые намекали после открытия дополнительного скелетного материала, извлеченного из типового местонахождения. Это было основано на наличии признаков, таких как твердое гнездо бедра (вертлужная впадина), и проксимального четвертого вертела на бедре, у которого также не было головы, обращенной внутрь, что типично для скелетов динозавра. Хотя он явно не был динозавром, он был предположительно истолкован как орнитодирский архозавр, все еще тесно связанный с динозаврами.
Открытие более полного материала с Мадагаскара подтолкнуло Флинна и его коллег к первой формальной классификации Azendohsaurus как не-динозавра, что в 2010 году благодаря подробному описанию его черепной анатомии и позволило уточнить его взаимосвязь. Вместо этого они признали, что это более базальный архозавроморф, не относящийся к кладе архозавров. Наряду с производными, подобными зауроподоморфным признакам, череп также имел многочисленные примитивные черты для архозавроморфов, включая нижнее височное дно с открытым дном, обширные небные зубы, отверстие шишковидной железы и отсутствие наружных нижнечелюстных или анорбитальных фенестр. Тем не менее, его точные отношения все еще оставались неизвестными за пределами позиции неопределенного неархозаврового архозавроморфа.
Признание Allokotosauria

{kind=link}
Azendohsaurus был впервые включен в филогенетический анализ триасовых архозавроморфов в 2015 году Несбиттом и его коллегами, используя всю новую информацию о черепе и скелете, а также обширную выборку различных видов триасовых архозавроморфов, где он был обнаружен как тесно связанный с другие загадочные травоядные триасовые рептилии, такие как трилофозавр и тератерпетон. Эта недавно признанная группа archosauromorphs была названа Allokotosauria, что означает «странные рептилии», за необычные качества рептилий, которые принадлежали к этой группе. Было установлено, что Azendohsaurus является родственным таксоном семейства Trilophosauridae, и был признан единственным членом своей семьи, Azendohsauridae, благодаря своей отличительной особенности даже среди других аллокотозавров. Аналогичный результат был получен в результате другого крупного анализа филогении архозавроморфов в 2016 году, проведенного Мартином Д. Эскуррой, который обнаружил монофилетическую аллокотозаврию, содержащую азендозавров и трилофозавров.
Аллокотозавры часто имеют специализированные челюсти и зубы, а также имеют ряд синапоморфий, которые включают в себя несколько обращений к более плезиоморфным (наследственным) признакам архозавроморфов, а также по меньшей мере два производных признака. Клада, как считается, хорошо поддерживается в этих анализах. Однако, хотя они тесно связаны, черепно-мозговые характеристики аллокотозавров резко различаются, и среди них у Azendohsaurus было характерно наличие поперечно сжатых зубчатых зубов по всей длине челюстей (в отличие от «клювовых» челюстей трилофосауридов). Azendohsaurus широко разделяет с другими особенностями azendohsaurids, такими как сливные ноздри, зубы в форме листьев и длинную шею, но сам Azendohsaurus отличается характерной бороздкой на внутренней поверхности верхней челюсти и зубных коронок, которые расширены над основанием.
В 2017 году Саради Сенгупта и его коллеги назвали еще одного крупного аллокотозавра из среднего триаса Индии по имени Shringasaurus indicus. Shringasaurus был очень похож на Azendohsaurus, и было обнаружено, что они тесно связаны, поддерживая существование Azendohsauridae как отличной семьи от трилофосауридов. Тот же самый анализ также обнаружил Pamelaria, еще одного длинношерстного архозавроморфа из Индии, в качестве базального азендохсаурида. Сходства между Pamelaria и Azendohsaurus были отмечены Несбиттом и его коллегами в 2015 году, в том числе слияниями ноздрей, зубчатых зубов и нижних шейных отделов позвоночника, но их анализ отдавал предпочтение позиции Allokotosauria, базальной к azendohsaurids. Анализ 2017 года также подтвердил тесную связь между A. laaroussii и A. madagaskarensis в пределах Azendohsauridae, усиливая их общее направление к роду Azendohsaurus. Анализ триасовых архозавроморфов, проведенный в 2018 году, не позволил восстановить аллокотозаврии, но все же обнаружил оба вида азендозавров в кладе азендохзаврид. Кладограмма ниже соответствует результатам Sengupta и коллег в 2017 году:
| Crocopoda |
| ||||||||||||||||||||||||||||||||||||||||||||||||
Эволюционное значение


{kind=link}
Сатурналия также имела конвергентное сходство с зауроподоморфами
Число сходящихся черт, общих для азендохзавра и зауроподоморф, удивительно велико, тем более что все общие черты интерпретируются как гомопластичные, что означает, что они развивались совершенно независимо друг от друга. Некоторые из общих адаптаций скелета между Azendohsaurus и sauropodomorphs ранее считались уникальными для sauropodomorphs. Однако конвергентная эволюция этих признаков у Azendohsaurus как адаптации к растительноядному образу жизни показывает, что они могут быть более широко распределены среди триасовых архозавроморфов и необязательно указывают на тесную связь с зауроподоморфами в ископаемых таксонах.
Характер сходства у Azendohsaurus необычен, так как они, по-видимому, возникли только в передней половине животного, в то время как раскидистые задние лапы и короткий хвост Azendohsaurus характерны для примитивных ранних архозавроморфов, очень отличающихся от столбчатых задних конечностей и длинных Хвост зауроподоморфов. Это также подчеркивает неравномерное распределение и приобретение типичных зауроподоморфных признаков у других растительноядных архозавроморфов.
Возраст Azendohsaurus также является значительным, так как он был примерно равным с самыми ранними известными зауроподоморфами из Южной Америки, такими как легкая двуногая сатурналия. Тем не менее, Azendohsaurus больше напоминает поздних норвежских зауроподоморфов, как по общей анатомии, так и по своему большему размеру тела. Это говорит о том, что azendohsaurids были первыми рептилиями, которые эволюционировали в качестве травоядных животных с высоким показателем роста в триасовых экосистемах, до развития более крупных зауроподоморфов, которые ранее, как предполагалось, были первыми травоядными животными с высоким показателем роста. Это также указывает на то, что конвергенция между Azendohsaurus и sauropodomorphs не происходила при тех же условиях окружающей среды, поскольку Azendohsaurus был частью начальной волны травоядных животных у archosauromorphs (наряду с rhynchosaurs, silesaurids и cynodonts) и большими sauropodomorphs с второй волной (травоядные псевдозухии).
Azendohsaurus также демонстрирует, что архозавроморфы играли роль крупных травоядных животных в триасовых экосистемах раньше, чем предполагалось. Ранее считалось, что в этих ролях преобладают крупные синапсиды, такие как дицинодонты, до облучения архозавров в позднем триасе, но Azendohsaurus предполагает, что более ранние архозавроморфы также могли конкурировать с синапсидными травоядными животными.
Палеобиология
Питание и диета
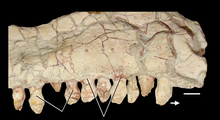
{kind=link}
Полностью развитые небные зубы предполагают, что он использовал их для кормления специализированным способом. Тем не менее, на небных зубах не было проведено никаких функциональных исследований, поэтому точно неизвестно, для чего они использовались, хотя их форма, сходная с краевыми зубами, позволяет предположить, что они использовались для обработки пищи. У крыловидного отростка у более молодого особи A. madagaskarensis меньше рядов небных зубов, которые меньше по размеру, чем у более крупных, зрелых особей, что указывает на то, что Azendohsaurus увеличивал как количество, так и размер своих небных зубов, когда он вырос во взрослую жизнь. У более молодых людей также было меньше зубных зубов, чем у взрослых, хотя разница была гораздо менее выраженной по сравнению с небными зубами (16 по сравнению с 17 зрелыми образцами).
Осанка
Положение тела, определенное для Azendohsaurus, аналогично таковому у ранних завроподоморфов - частично он ходил на двух ногах, частично - на четырёх. Задние конечности были интерпретированы как полностью вытянутые наружу от тела, с бедром, выпрямленным прямо, а задние лапы перпендикулярны поверхности ниже колена. Тем не менее, передние конечности и плечевой пояс предполагают, что передняя часть тела держалась более вертикально, чем задняя, с частично направленной вниз и плечевой костью, более подходящей для удержания частично выпрямленного положения, и была похожа по форме на таковую у завроподоморфов. Эта необычная комбинация предполагает, что Azendohsaurus стоял с поднятой над землёй передней частью тела, что в сочетании с его длинной изогнутой шеей и маленькой головой позволяло ему относительно высоко подниматься над землей, в отличие от современных ему низколобых ринхозавров и цинодонтов. Адаптация к просмотру с высоты могла бы объяснить сближение Azendohsaurus и sauropodomorphs, приобретая подобные черты шеи, передних конечностей и позвоночника, чтобы действовать в подобных нишах.
Палеопатологии


{kind=link}
Несмотря на множество образцов, которые были исследованы, у A. madagaskarensis была зарегистрирована только одна патология. Образец UA 7-16-99-620, один из трех сохранившихся межключичных, был деформирован так, что длинный задний отросток был резко согнут вправо, по сравнению с нормальными прямыми задними отростками двух других межключичных.
Метаболизм и рост
В 2019 г. из плечевой, бедренной и большеберцовой костей были вырезаны тонкие срезы образцов, приписанных A. laaroussii, для гистологического исследования микроскопической структуры кости, чтобы попытаться определить скорость роста у Azendohsaurus. Было обнаружено, что плотность сосудов (плотность кровеносных сосудов в костной ткани) во всех трех костях конечностей сопоставима с таковой у быстрорастущих птиц и млекопитающих, а также идентифицированы типы костной ткани, в частности энергопотребляющая фиброламеллярная костная ткань, были интерпретированы как указывающие на высокий уровень метаболизма в покое, который наблюдается у живых птиц и млекопитающих. Тогда было сделано заключение, что, подобно птицам и млекопитающим, Azendohsaurus также был бы эндотермальным или теплокровным. Высокие показатели метаболизма в покое, сходные с таковыми у Azendohsaurus, были выявлены у других более производных архозавроморфов (таких как Prolacerta), и анализы предположили, что эндотермия, возможно, в то время имела наследственное происхождение у архозавроморфов еще до появления аллокотозавров. Это говорит о том, что тогда Azendohsaurus мог быть наследственно эндотермальным. Напротив, ранее было обнаружено, что родственный аллокотозавр Trilophosaurus не имеет какой-либо фиброламеллярной костной ткани в костях конечностей, и поэтому предполагалось, что он рос медленно.
Палеоэкология
Хотя два вида Azendohsaurus известны из разных мест: в Северной Африке и на Мадагаскаре, во время среднего и позднего триаса эти регионы были соединены как часть суперконтинента Пангея. Из-за этого эти два региона разделяют в значительной степени сходные фауны, а также разделяют некоторые с другими регионами земного шара в то время. Например, цинодонты на Мадагаскаре похожи на те, что встречаются также в Южной Америке, а марокканские темноспондилы могут быть связаны с таковыми в восточной части Северной Америки. В это время климат был жарким и сухим, но есть свидетельства того, что во время карна начало выпадать больше осадков, что прерывает тенденцию к увеличению засушливости и создает более влажную среду по всему земному шару.