![]()
Птериготиоиды (лат. Pterygotioidea) — надсемейство крупных эвриптерин, обитавших на территории Евразии и обеих Америк с силура по девон, 442–372.2 млн лет назад. Почти поголовно были апексхищниками.
История
Раннее изучение
Первые останки птериготиоидов принадлежали птериготу, которого первооткрыватель Луи Агассис посчитал в 1839 году большой рыбой. Собственно, "Pterygotus" и означает "крылатая рыба". Через пять лет Агассис признал ошибку первоначальной классификации и определил птеригота к членистоногим на основе новых образцов, найденных в красных песчаниках Шотландии.[1]

Slimonia Page, 1856
Вскоре начали находить и другие виды птериготов. Так, в 1856 в Шотландии нашли другой вид — "Pterygotus acuminata", отличающийся от оригинального птеригота формой панциря и отсутствие хелицеровых когтей. Вскоре "P. acuminata" был выделен в собственный род — Slimonia acuminata. В 1859 году были описаны птериготы P. osborni и P. marcophthalmus, позже синонимизированные. В 1872 году в Богемии нашли P. bohemicus, а через три года в Нью-Йорке обнаружили два вида - P. buffaloensis и P. cummingsi.
В 1903 году на основе найденных в Вернонской формации в Питтсфорде окаменелостей была выделена хьюджмиллерия. Описавший её Клифтон Дж. Сарл предположил, что она является промежуточным звеном между базальным эвриптером и птериготом, однако не приписал хьюджмиллерию к какому-либо семейству.
Создание семейства

Acutiramus Ruedemann, 1935
После синонимизации P. buffaloensis и P. cummingsi в 1912 году, Кларк и Рюдеман отметили, что P. macrophthalmus отличается от остальных видов птеригота. Хотя он был тесно связан с P. anglicus, эти два вида можно легко отличить по нескольким характеристикам, в частности, по форме охотничьих придатков и их направлению. Особенности придатков интерпретировали как сходство с P. osiliensis, вида с двойным тельсоном. Кроме того, Кларк и Рюдеман отметили, что P. macrophthalmus имеет свободную ветвь нижней челюсти, имеющую переходные признаки P. cummingsi и остальных птериготов.
Вскоре в роде Pterygotus скопилось просто огромное число видов и результатом этого стало создание в 1912 году Рудольфом Рюдеманом и Джоном Кларком семейства птериготиды, включавшее тогда роды птеригот, хьюджмиллерия, слимония и хастимима. А птеригот, к тому же, был разделён на несколько подродов: Pterygotus (Erettopterus), Pterygotus (Curviramus) и Pterygotus (Acutiramus). Эреттоптера вскоре выделили в собственный род, однако в 1951 году Эрик Кьеллесвиг-Ваеринг оставил в составе птериготид только птеригота, выделив слимонию, хастимиму, хьюджимллерию и недавно открытого гроссоптера в семейство хьюджиллериды.

Erettopterus Salter in Huxley & Salter, 1859
Лейф Стёрмер в 1955 году предположил, что птериготиоиды являются кладой в составе надсемейства эвриптерасы. Однако в 1962 году Нестер Иванович Новожилов переклассифицировал эвриптерас до подотряда, синонимизировав с эвриптеринами. Птериготиоидов же Новожилов определил как надсмейство, содержащее птериготид и хьюджмиллерид и недавно выделенных слимонид, содержащих тогда только слимонию. Новожилов также переклассифицировал хастимиму и гроссоптера в качестве миктероптерида и вэрингоптерида соответственно, оставив в семействе хьюджмиллериды только хьюджмиллерию.
Последующее изучение


Jaekelopterus Waterston, 1964
В 1964 году был выделен крупнейший род птериготиоидов — йекелоптер. Решение о создании нового таксона было сделано на основе различий в строении полового придатка. Некоторые учёные даже предложили выделить его в семейство йекелоптериды, однако найти серьёзных различий между йекелоптером и остальным птериготидами не удалось. В 1974 году Стёрмер переклассифицировал акутирама и трункатирама в качестве самостоятельных родов, однако второго вскоре синонимизировали с эреттоптером.[2]

Herefordopterus Tetlie, 2006
В 1986 году Пол Селден исследовал останки загадочного членистоного некрогаммара и пришёл к выводу, что останки принадлежат крупному птериготиоиду, скорее всего, либо эреттоптеру E. marstoni, либо птериготу P. arcuatus. Оба птериготида найдены в той же местности, что и некрогаммар, однако из-за фрагментарности последнего нельзя определить его к кому-то конкретно. Сейчас некрогаммар считается неопределённым птериготидом.
После установления некоторых схожих черт между слимонией и ракоскорпионом сальтероптером, Виктор Толлертон определили последнего к слимонидам в 1989.[3]
В 2006 году один из видов хьюджмиллерии, H. banksii, был выделен Эриком Тетли в самостоятельный род - герефордоптер. При наличии двух родов в семействе скоро были выделены отличительные признаки хьюджмиллерид, вроде остистых придатков.[4]
В 2009 году птеригот Pterygotus ventricosus был выделен в собственный род чуркоптер. Это самый базальный член всех птериготид, доказавший, что именно слимониды являются ближайшими родственниками птериготид, а не хьюджмиллерии или герефордоптеры, как считалось ранее.
Описание

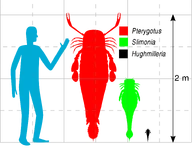
Сравнение размеров птеригота, слимонии и хьюджмиллерии
Птериготиоиды обитали с раннего силура по поздний девон. Отличить их от других ракоскорпионов можно по расположению сложных глаз - они находятся по краям или вблизи карапакса - заднего щитка просомы. Размеры птериготоид варьировались от 6 сантиметров (хьюджмиллерия Hughmilleria wangi) до 2,5-2,6 метров (йекелоптер Jaekelopterus rhenaniae).
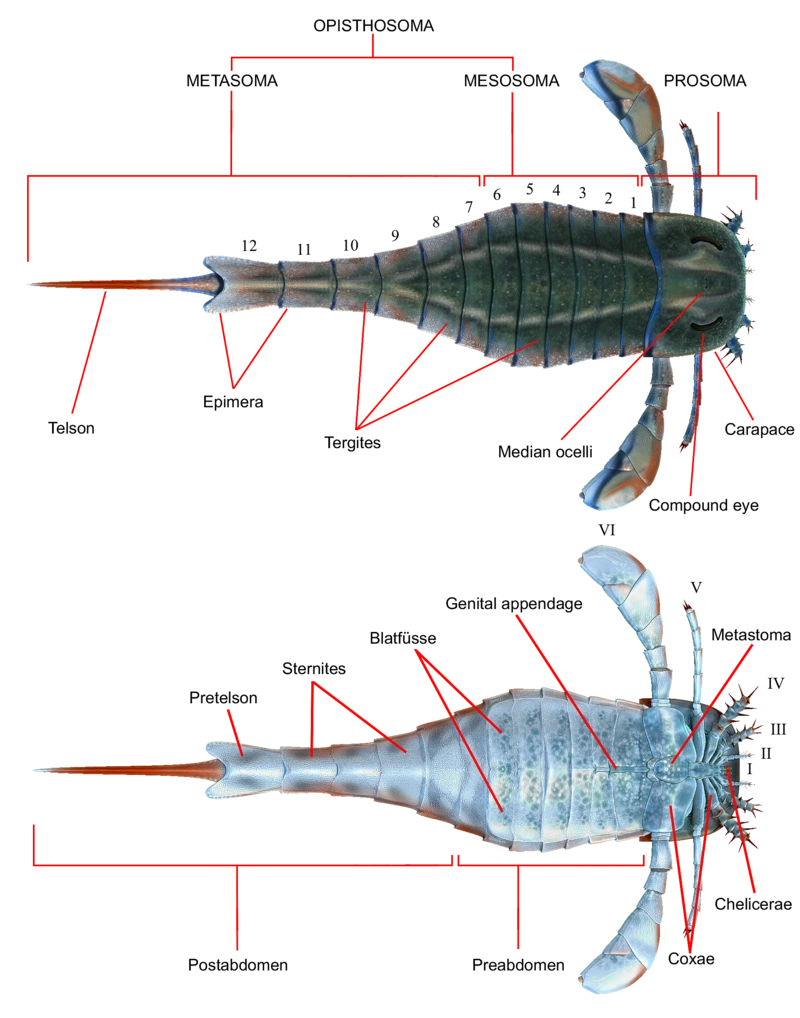

Строение эвриптера на примере аделофталмоида
Как и все членистоногие, птериготы имели сегментированное тело и особые придатки - хелицеры. У членов семейства птериготиды панцирь был покрыт чешуйчатым орнаментом, в отличии от слимонид и хьюджмиллерид.
Тело эвриптерид делилось на две части - просому, голову, и опистосому, туловище.
Как и их дальние родственники паукообразные, птериготиды имели хелицеры — особые придатки, использующиеся для питания. Эта функция сохранилась у хьюджмиллерид и слимонид. Развитые и узкоспециализированные птериготиды использовали хелицеры как орудие охоты, а не питания. На них находились особые когти — хелы. Хелицеры крепились к просоме и имели форму клешней. У слимонид и птериготид они были лишены шипов и были тонкими и заострённые. Хелицеры примитивных хьюджмиллерид обладали шипами. Интересно, что у некоторых птериготид, вроде акутирама, хелицеры не подходили для охоты на некрупных животных. Возможно, обладатели таких клешней были падальщиками или вовсе растительноядными.
Тельсон производных птериготид и слимонид был расширенными и уплощёным, часто оканчивался шипом. Хьюджмиллериды же имели тельсон в виде копья или шипа, чем напоминали более примитивных ракоскорпионов вроде эвриптера. Интересно, что хьюджмиллерид герефордоптер имел киль и особый орнамент по краям тельсона, что роднит его с птериготидами.


Птеригот, эвриптер и стилонур от Йосефа Смита
Отличительной чертой хьюджмиллерид было наличие шипов на второй-пятой паре плавательных ног с узким седьмым и восьмым элементом. Девятый элемент имел обтекаемую форму. Передняя часть панциря хьюджмиллерид была более широкой, чем задняя.
Отличительные черты слимонид неясны, так как сальтероптер известен только по частям тельсона и метастому. Слимония же известна по многочисленным останкам, в том числе и полным окаменевшим панцирям. Скорее всего, отличия слимонид от птериготид и хьюджмиллерид заключаются в квадратной просоме, размещению глаз, небольших хелицерах с развитыми когтями и форме панциря.
Систематика
Птериготиоиды относятся к отряду эвриптеры в составе подотряда эвриптерины. Инфраотряд диплоперкулаты, к которому относятся птериготиоиды, содержит наиболее производных и развитых ракоскорпионов — аделофталмид, птериготиоид, карциносоматиод и вэрингоптерид. Отличительной чертой всех этих эвриптеров является генитальный оперкулум (структура, содержащая генитальный придаток), состоящий из двух соединённых сегментов.
Птериготиоиды являются наиболее производными членами диплоперкулат и насчитывают 50 видов, являясь самой многочисленной группой. Это примерно одна пятая от всего разнообразия эвриптеров. Ближайшая группа — монофилейное надсемейство аделофталмоиды с единственным семейством аделофталмиды, на которых приходится второе место по разнообразию (40 видов). Ниже дана кладограмма инфраотряда диплоперкулаты:
| Diploperculata |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||


Хотя птериготиоиды считаются наиболее производной группой, ведутся споры о степени родства хьюджмиллерид и слимонид с птериготидами, а также о том, какое семейство является более примитивным. Их удалось разрешить благодаря открытию чуркоптера, сочетающего в себе признаки как слимонид, так и более развитых птериготид. Таким образом, слимониды являются сестринским таксоном для птериготид.


Сравнение размеров хьюджмиллерий
Отсутствие орнамента на тельсоне хьюджмиллерии и наличие некоторых общих с аделофталмидами черт, вроде треугольного края просомы, делает её самым базальным членом птериготоид. Кроме того, хьюджмиллерия имеет больше гнатобазовых зубов, чем любой другой птериготиоид, в том числе и её родственник герефордоптер.
Ниже дана кладограмма Эрика О. Тетли 2007 года, показывающая отношения птериготид: в то время как слимониды с птериготидами являются монофилейными группами, хьюджмиллериды являются парафилейногруппой. Из-за этого их нельзя расценивать всерьёз как семейство, но хьюджмиллериды регулярно появляются в филогенетических анализах.
| Pterygotioidea |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Появление и распространение


Наиболее часто эвриптеров находят на Скандинавском полуострове, в Восточной Европе, Англии и США. В силуре-девоне все эти регионы составляли материки Балтику (Скандинавия и Восточная Европа), Авалонию (Германия, Великобритания и часть Северной Америки) и Лаврентия (большая часть Северной Америки). Именно в этих регионах чаще всего и находят птериготиоид. Кроме того, останки некоторых членов этого надсемейства находят и в местах, где не было других эвриптеров — в Австралии, Марокко, Ливии, Флориде, Саудовской Аравии, Китае, Франции, Южной Америке, Сибири и Чехии, что указывает на значительное распространение данной группы в период её существования.
Рассвет птериготиоидов пришёлся на средний силур-средний девон, причём они были единственными ракоскорпионами, добравшимися даже до южного материка Гондваны.


Реконструкция аделофталма - типичного представителя аделофталмидов
Самые ранние птериготиоиды были некрупными и относятся к роду хьюджмиллерия. Датируются Лландоверской эпохой силурийского периода и встречаются на территории Лаврентии, Британии и южного Китая. Такой широкий ареал усложняет поиски предка птериготиоидов, однако известно, что самыми близкими родственниками данной группы было семейство аделофталмиды. Первые члены этой группы появились в окрестностях Лаврентии, так что искать предков и самых ранних птериготиоидов стоит именно там.
Неясно, могли ли птериготиоиды мигрировать по дну океана, или же виды, обнаруженные за пределом Лаврентии, Балтики и Авалонии, являются единичными изолированными представителями. Однако аделофталмиды и птериготоиды были превосходными пловцами, так что они запросто могли пересекать межконтинентальные моря.
Палеоэкология и палеобиология
Хьюджмиллериды


Акутирам, стилонур, эусакрана, эвриптер и долихоптер. Хьюджмиллерию можно заметить в нижнем правом углу
Окаменелости хьюджмиллерид находят с лландоверской по ладловской эпохи силура США, Китая и Англии. В то время как хьюджмиллерия обитала в солоноватых водах, герефордоптер предпочитал мене солёную бентосную приливно-отливную зону побережий. Возможно, они могли напоминать своим поведением крабов, спокойно передвигаясь по берегам и дыша атмосферным воздухом.
В зоне Питтсфордских сланцев удалось обнаружить огромную общину ракоскорпионов: кроме хьюджмиллерии там обитали эреттоптер E. osiliensis, эвриптер E. pittsfordensis, миксоптер M. multispinosus и карциносома C. spiniferus. А в графстве Херефордшир, где обнаружили герефордоптера, удалось найти огромное число других эвриптеридов: эреттоптер E. gigas, эвриптер E. cephalaspis, нанахьюмиллерия N. pygmaea, марсупиптер M. sculpturatus, сальтероптер S. abbreviatus и слимония S. stylops.
Слимониды

Симония и анаспид
Отличительной чертой слимонид можно считать большой уплощённый тельсон, который они разделяют с птериготидами, гиббертоптером и хастимимой. Однако у последней он разделился на двое в процессе эволюции.
Назначение таких тельсонов до конца не выяснено и спорно, однако их функция у слимонии и её родственников могла заключаться в передвижении. Считается, что птериготиды, перемещая весь нижний щит, а тельсон и опистосома в таком случае обеспечивали бы волнообразные пропульсивные движения. Плавательные ноги тогда никак не использовались, чем птериготиды отличались бы от остальных ракоскорпионов.
Однако находки противоречат данной гипотеза. Панцири эвриптеров были жёсткими с обеих сторон, а отпечатки не сохранили никаких механизмов, обеспечивающих гибкость данной конструкции. К тому же, любое сгибание тела требует мышечных сокращений, и, опять же, ни одна окаменелость птериготоид не показывает крупных аподем (внутренних гребней экзоскелета, к которым крепятся мышцы) или мышечных рубцов, указывающих на наличие сильных описостомальных мышц. Вместо этого обнаружены признаки передвижения с помошью последней пары плавательных ног, что характерно для ракоскорпионов.
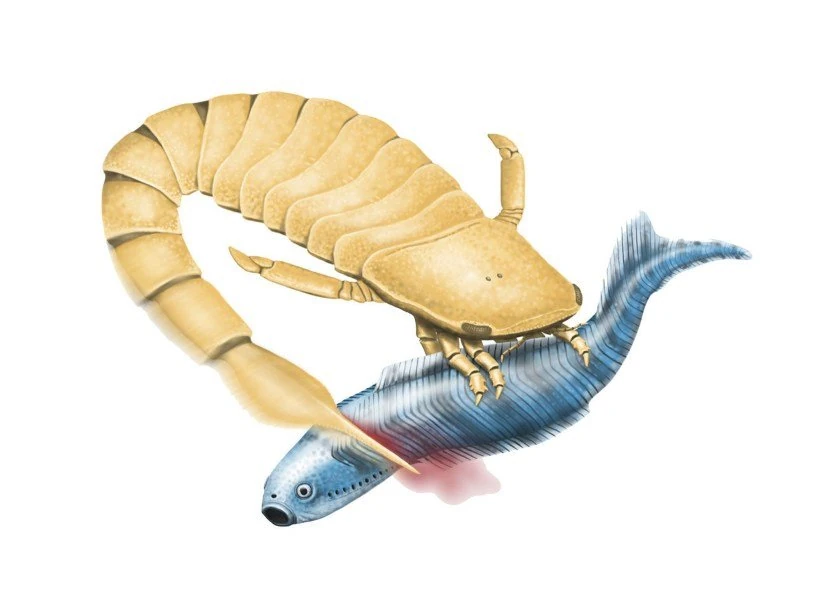
Ошибочная реконструкция охоты слимонии
Биомеханические исследования тельсонов и постабдоминальных сегментов тесно связанных эвриптерид, особенно тех из семейства птериготид, показали, что тело было очень жестким, а сплющенный тельсон, вероятно, служил в качестве руля, позволяя животным быть способными к быстрым поворотам, которые они делали, преследуя добычу. Это противоречит предыдущим гипотезам, в которых тельсоны рассматривались как средство наземного передвижения. Возможно, что хвост использовался как оружие, двигающимся только вправо-влево.
Однако, позже теорию "оружия" опровергли. Хотя первоначальное исследование не показало признаков расчленения, скорее всего, оно ошибочно. Видно, что тергиты 8 и 10 неестественно перекрывают друг друга, кроме того, этот образец - сброшенный панцирь, результат линьки, а не туша. То есть, поза окаменелости никак не может указать на какую-то позицию при жизни существа. Кроме того, хвостовой киль создавал значительное сопротивление, что тоже мешало бы удару. Да и зубцы на тельсоне, скорее всего, служили для прикрепления щетинок.
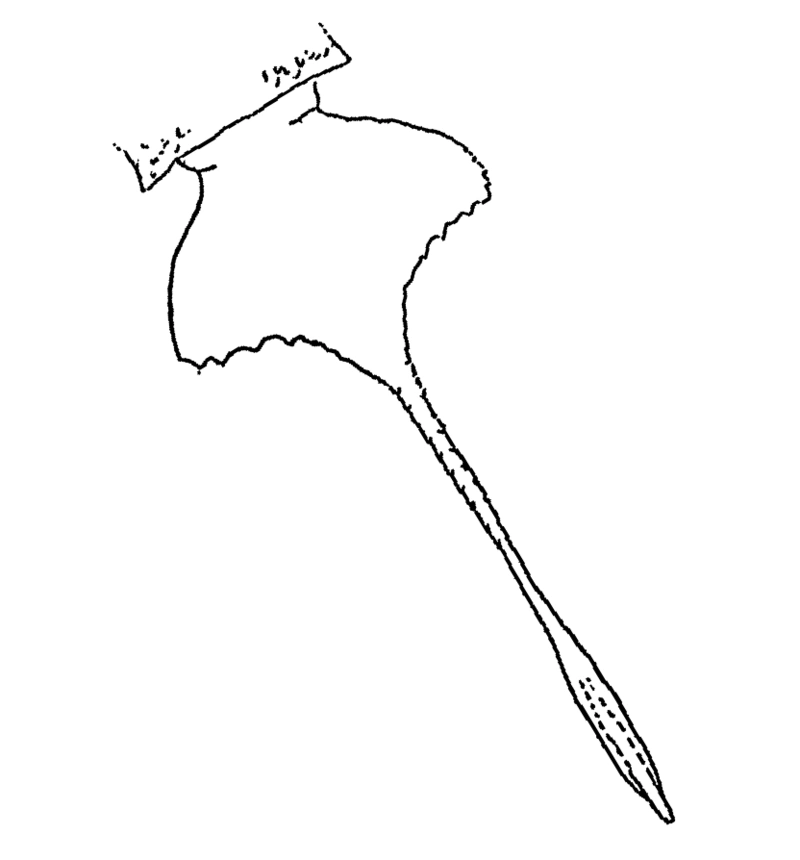
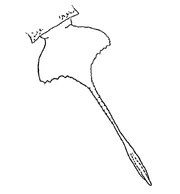
Тельсон сальтероптера
Тельсон сальтероптера отличается от тельсона слимонии, его предназначение тоже неясно. Возможно, он исполнял роль балансира при передвижении по дну или песку. Уплощённая часть тельсона тригональна и меньше, чем у слимонии, однако хвостовой шип у сальтероптера гораздо длиннее и имеет несколько бугорков. На передней части тельсона находится странное образование, уникальное среди ракоскорпионов.
Птериготиды
Ранее считалось, что все птериготиды — активные хищники, однако исследование хелицеров и глаз показали, что их можно разделить на несколько строго специализированных групп. Для морфологии эвриптерид и птериготиоид в частности важна острота зрения: по ней можно узнать условия, в которых жил ракоскорпион. Остроту и ясность зрения у членистоногих можно узнать путём определения числа линз в сетчатых глазах межкомматидиального угла. Этот угол особенно важен для различения разных по экологической роли членистоногих. У современных членистоногих хищников он очень низок.


Эреттоптеры охотятся на бесчелюстных
Несмотря на морфологическое сходства, экологические ниши птериготид очень сильно различались. Так, по остроте зрения эреттоптер был схож со слимонией. Эти двое имели зрение лучше, чем у производного акутирама, но хуже, чем у доминантных хищников птеригота и йекелоптера. А строение хелицеров эреттоптера предполагает, что это был неспециализированный охотник или падальщик. Хелицеровые когти у эреттоптера, как и у других птериготид, увеличены, однако лишены зубчиков. Скорее всего, они использовались только для хватания добычи. Хотя количество линз в глазах эреттоптера близко к более поздних птериготидам, он не был активным или узкоспециализированным охотником, как йекелоптер или птеригот.
Острота зрения акутирама была очень низкой, что не подходит под описание активных хищников-птериготид. Число линз межкомматидиального угла менялось во время взросления, и острота зрения у взрослых особей была меньше, чем у молодых, что отличает его от других птериготид, вроде йекелоптера. По этому же признаку можно понять, что акутирам занимал другую экологическую нишу, чем его собратья-птериготиды: он мог быть падальщиком или охотиться из засады. Возможно также, что эта разница в остроте зрения обуславливалась ночным образом жизни акутирама.


Хелицеровые когти
Хелицеры птериготид служили для той же цели, что и клешни ракообразных или современных паукообразных. Основная функция данного органа состояла в захвате, разрезании или раздроблении пищи на более мелкие удобные кусочки и защите хозяина от врагов.
Хотя большинство эвриптеров имело простые клешни, у птериготид хелицеры показывают уникальное приспособление для захвата и удержания добычи. Кроме того, они были больше, чем у какого-либо другого представителя отряда.
Хелицеры состояли из нескольких основных элементов и трёх-пяти суставов в зависимости от вида. Раньше считалось, что у птериготиоидов было три сустава — один большой базальный и два маленьких дистальных. Однако последние исследования неполных образцов актирама и эреттоптера показали, что, скорее всего, хелицеры состояли из четырёх суставов. Это обеспечивало подвижность клешней, а если бы придатки состояли из трёх суставов, то хелицеровые когти находились бы на самом кончике клешней, становясь тем самым бесполезными.
Первый сустав хелицеров, находившийся на месте крепления к эпистоме, обеспечивал перемещение клешней в практически любом направлении, из-за чего исследователи пришли к выводу, что главной задачей хелицер у птериготид была не защита, а захват и разрезание добычи для более удобного поедания. Рот эвриптера не предназначен для захвата крупных кусков мяса, а ходячие ноги были бесполезны для разрезания, поэтому некоторые прогрессивные ракоскорпионы изменили строения своих хелицеров для более удобного захвата и раздробления пищи. Один из когтей поддерживал тело жертвы, в то время как другой отрывал подходящий кусок плоти и переносил его ко рту.
Хелицеры, как было сказано выше, различались от образа жизни. У птеригота и йекелоптера они могли быть не только орудием захвата, но ещё и колющим и дробящим орудием. У акутирама хелицеры хоть и были большими, но выполняли только режущую и удерживающую функции.


Как птеригот, так и акутирам, имели высокую остроту зрения, что было подтверждено низким значением межкомматидиального угла и большим количеством линз в сложных глазах. Хелицеры этих двух ракоскорпионов были большими и крепкими, обладали изогнутыми когтями разной длины и размера, что соответствует сильным колющим и хватательным движеиям. Скорее всего, птеригот и акутирам были активными хищниками.
Тельсон


Эреттоптер обладал редкой разновидностью тельсона
Тельсон птериготид большой и уплощённый, как у слимонид и производных гиббертоптера и хастимимы. Имеет плоскую форму с приподнятым килем. У некоторых представителей семейства задний край тельсона имеет несколько шипов. Среди таких стоит отметить птеригота и акутирама. У эреттоптера тельсон имеет необычную раздвоенную форму.
Долгое время велись споры вокруг предназначения этих органов. Так, Кьеллесвиг-Веринг в 1964 году сравнил тельсон птериготиоида и хвостовую двуустку кита, предполагая использование тельсона в качестве дополнительного органа волнообразного передвижения. Однако, как было сказано выше, панцирь эвриптерид был слшиком жёстким и нет никаких доказательств того, что он мог изгибаться при передвижении.
Альтернативную теорию предложил К. Д. Уотерстон в 1979 году. Он предположил, что тельсон использовался в качестве руля при плавании. Расчёты и гипсовые модели птериготид позволили Плотнику в 1988 году доказать, что члены данного семейства были весьма проворными охотниками, способными совершать быстрые повороты при преследовании добычи.
Гигантизм


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
_(5977251311).jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Среди птериготидов известны случаи гигантизма. Так, йекелоптер и его близкий родственник акутирам были самыми большими ракоскорпионами, достигая в длину 2,5 и 2,1 метра соответственно. Недавно было предложено несколько факторов, способствующих гигантизму этих видов.


{kind=link}
Возможны ли ракоскорпионы крупнее, чем йекелоптер?
Факторы, как например, дыхание, свойства экзоскелета, могут ограничивать размер членистоногих. Большая часть сохранившихся останков сегментов очень тонкая. Образование брюшных сегментов было очень тонкое, и можно предположить, что конструкция экзоскелета животного была очень легкой, что не ограничивало животное в передвижении. Такие приспособления были и у других палеозойских членистоногих, например, у артроплевры, и таким образом, облегченный экзоскелет меньше ограничивает размер животного. Не смотря на то, что ракоскорпионы были самыми длинными членистоногими за историю Земли, из-за облегчённого экзоскелета они не были самыми тяжёлыми членистоногими.