![]()
Цааган (лат. Tsaagan) — род плотоядных динозавров семейства дромеозаврид из формации Djadokhta, позднего мелового периода Монголии. Ископаемые останки Tsaagan были обнаружены в 1996 году и поначалу идентифицированы как принадлежавшие велоцираптору. После рентгеновского сканирования в мае 1998 года был сделан вывод о принадлежности останков новому роду. В декабре 2006 года типовой вид был описан группой учёных под руководством Марка Норелла. Название вида, Tsaagan mangas, следует рассматривать как единое целое; словосочетание переводится с монгольского как «белый монстр», несмотря на случайную ошибку в слове цагаан.
Описание

{kind=link}
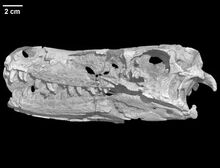
Образец голотипа, IGM 100/1015, был найден недалеко от Ксанаду, провинция Умнеговь, в слоях формации Djadokhta, которая датируется кампанским ярусом мелового периода (около 75 миллионов лет назад). Образец состоит из хорошо сохранившегося черепа и серии из десяти шейных позвонков, а также поврежденной левой части плечевого пояса. Это единственный найденный экземпляр Tsaagan. Он принадлежал взрослой особи.
Череп голотипа имел длину 201 миллиметр. Морда длинная, составляет 59% от длины черепа, но довольно высока спереди, так что голова имеет полый верхний профиль, но без резких изгибов, как у Velociraptor. Передняя челюсть имеет короткую заднюю ветвь по направлению к верхней челюсти. Ветвь к носовой кости изящно построена. Верхняя челюсть, длиной 103 миллиметра, крепкая, но с небольшой боковой полостью, fossa antorbitalis, которая в основном заполнена большим отверстием черепа, fenestra antorbitalis и относительно большим fenestra maxillaris спереди, который касается края ямки. Перед ним снова небольшое щелевидное отверстие промаксиллярного типа. Ноздри овальной формы и большого размера, но не вызывает выпуклости на голове; таким образом, верхняя часть черепа образует плавную элегантную кривую, по которой образец можно сразу отличить от окаменелостей велоцираптора. Однако авторы считают, что это происходит главным образом из-за того, что последние гораздо сильнее сжаты и что хорошо известный «курносый нос» велоцираптора слишком сильно задействован в большинстве реконструкций. Носовая кость простирается назад за передний край глазницы. Там он касается Т-образной слезной кости, которая не выходит за пределы черепа. Небольшой рог перед глазом, как у многих родственников, здесь отсутствует, что добавляет общей плавности. Компьютерная томография показала, что внутри слезной кости имеется слезный канал, а на переднем крае с глубокими бороздками, который сразу образует задний край анторбитального отверстия, по-видимому, находится какая-то железа.
{kind=link}
Надбровные дуги образуют верхнюю часть прямоугольных глазниц по бокам и там довольно шероховаты. Описатели сомневаются, было ли это началом надбровной кости или глазного дна; возможно, он образовывал прикрепление декоративного рогового ободка над глазом. Лобная кость не имеет глубокого углубления на переднем крае впадины вокруг верхнего височного окна. Он также не имеет широкого выступа в посторбитальную область и, следовательно, лишен типичной Т-образной формы дромеозаврид; он несколько пневматизирован. Сзади надбровные кости касаются теменных костей, составляющих верхнюю часть черепной коробки. На своей средней линии они образуют довольно высокий центральный гребень; в их задней части исключительно высокий поперечный гребень, указывающий на сильную мускулатуру шеи.
Треугольная высокая скула составляет большую часть нижнего края глазницы. Он очень гладкий, без выемки на боковой поверхности спереди и гребня сзади. Внутри массивный, без воздушных полостей. Задний край имеет полый профиль, благодаря чему нижнее окно для сна имеет форму почки. Восходящая ветвь касается чешуйчатой кости, так что посторбитальная ветвь не влияет на верхний край этого окна для сна; этот аутапоморфизм уникален среди Deinonychosauria, но эта характеристика в основном вызвана тем фактом, что рассматриваемое fenestra infratemporalis очень маленькое, а не потому, что скула теперь имеет совершенно другую форму. Сама посторбиталь маленькая, треугольная и плоская. Квадратично-скуловая кость имеет короткую восходящую ветвь, которая составляет только половину заднего края нижнего окна для сна. Верхняя половина образована нисходящей ветвью четырехлучевой крупной чешуи. Эта кость имеет внутреннюю полость по направлению к внутренней части черепа, нижний край которой образует горизонтальный гребень; он шире, чем когда-либо был обнаружен у любого другого теропод, однако у большинства видов эта часть не сохранилась в окаменелостях. На заднем крае черепа квадратная клетка имеет небольшое, по-видимому, пневматическое отверстие, но внутри она сплошная, как показано на сканировании.


{kind=link}
На заднем крае черепной коробки поперечный гребень образует широкий воротник, задняя часть которого, образованная затылочной костью, спускается посередине вниз к овальному затылку. Слева и справа от этого склона лежат еще более глубокие впадины, внешние стенки которых, все еще образованные теменными костями, чуть выпуклые в крайние стороны воротника черепа. Длинные крыловидные праокципитальные отростки выступают по диагонали с обеих сторон затылка; их концы слегка свисают. Ниже затылка находится затылочный мыщелок, соединение с шейными позвонками; под ним слева и справа два меньших изогнутых вверх выступа, базиптеригоиды, которые текут боком в черепную коробку. Базиптеригоиды исключительно длинные, крупнее любых других известных дромеозаврид. Сбоку от черепной коробки верхняя наклонная направленная вверх часть, которая образует внутреннюю стенку верхнего окна для сна, отделена от нижней четким выступом. В нижней части имеется множество сложных отверстий для нервов, разделенных гребнями, включая небольшое барабанное углубление. Полукружные каналы в ухе свободны
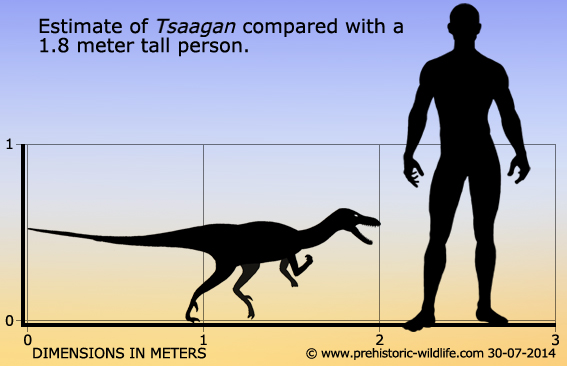
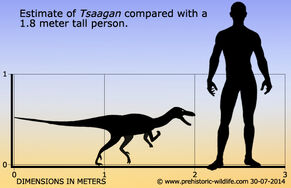
Tsaagan был дромеозавридом среднего размера. В 2010 году Грегори С. Пол оценил его длину в два метра, а вес в пятнадцать килограммов. Его череп по внешнему виду напоминает череп велоцираптора, но отличается от него многими деталями. Его вершина более крепкая и ровная; уникальные черты, полученные в результате аутапоморфии, включают в себя длинные отростки в затылочной части черепа и скуловую кость, примыкающую к чешуйчатой кости.
Tsaagan был отнесён к семейству Dromaeosauridae. Сделанный Нореллом и его коллегами кладистический анализ показал, что динозавр является членом подсемейства Velociraptorinae. Анализ 2010 года выявил его тесное родство с линьхэраптором; впоследствии Сентер (2011) и Тёрнер с коллегами (2012) утверждали, что Linheraptor exquisitus фактически является младшим синонимом Tsaagan mangas.
Останки Tsaagan являются единственными известными образцами из области Ukhaa Tolgod, хотя другой дромеозаврид, велоцираптор, был найден в той же формации. Животными, делившими с этими дромеозавридами среду обитания, были протоцератопс, шувууйя, маленькое млекопитающее залямбдалестес, многобугорчатое млекопитающее Kryptobaatar, а также несколько ящериц и два пока не описанных вида троодонтид и дромеозаврид.
Классификация
Приведённая ниже кладограмма отражает результаты анализа, проведённого в 2015 году группой палеонтологов под руководством Роберта ДеПалмы на основе данных, полученных из Theropod Working Group.
| Дромеозавриды |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Кладограмма 2020 года:
| Эудромеозавры |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Литература
- Norell, M.A.; Clark, J.M.; Turner, A.H.; Makovicky, P.J.; Barsbold, R.; Rowe, T. (2006). "A new dromaeosaurid theropod from Ukhaa Tolgod (Ömnögov, Mongolia)". American Museum Novitates. 3545: 1–51. doi:10.1206/0003-0082(2006)3545[1:ANDTFU]2.0.CO;2. hdl:2246/5823.
- Paul, G.S., 2010, The Princeton Field Guide to Dinosaurs, Princeton University Press p. 137
- Xu, X., Choinere, J., Pittman, M., Tan, Q., Xiao, D., Li, Z., Tan, L., Clark, J., Norell, M., Hone, D.W.E. and Sullivan, C. (19 March 2010). "A new dromaeosaurid (Dinosauria: Theropoda) from the Upper Cretaceous Wulansuhai Formation of Inner Mongolia, China" (PDF). Zootaxa (2403): 1–9. Retrieved 2010-03-19.
- Senter, Phil (2011). "Using creation science to demonstrate evolution 2: morphological continuity within Dinosauria (supporting information)". Journal of Evolutionary Biology. 24 (10): 2197–2216. doi:10.1111/j.1420-9101.2011.02349.x. PMID 21726330.
- Alan Hamilton Turner, Peter J. Makovicky and Mark Norell (2012). "A review of dromaeosaurid systematics and paravian phylogeny". Bulletin of the American Museum of Natural History. 371: 1–206. doi:10.1206/748.1. hdl:2246/6352. S2CID 83572446.
- Xu Xing; Michael Pittman; Corwin Sullivan; Jonah N. Choiniere; Qing Wei Tan; James M. Clark; Mark A. Norell; Wang Shuo (2015). "The taxonomic status of the Late Cretaceous dromaeosaurid Linheraptor exquisitus and its implications for dromaeosaurid systematics". Vertebrata PalAsiatica. 53 (1): 29–62.
- Currie, P. J.; Evans, D. C. (2019). "Cranial Anatomy of New Specimens of Saurornitholestes langstoni (Dinosauria, Theropoda, Dromaeosauridae) from the Dinosaur Park Formation (Campanian) of Alberta". The Anatomical Record. 303 (4): 691–715. doi:10.1002/ar.24241. PMID 31497925.
Источники
- https://en.wikipedia.org/wiki/Tsaagan
- http://a-dinosaur-a-day.com/post/119954660885/tsaagan-mangas
- https://ru.wikipedia.org/wiki/Tsaagan
- http://www.prehistoric-wildlife.com/species/t/tsaagan.html
- http://www.dinosaurfact.net/Cretaceous/TsaaganMangas.php
Галерея
")
")
")
")
")
")