 |
Данная статья находится в работе! |
 |
Конкавенатор (лат. Concavenator) — горбатый ящер из Испании, родственник акрокантозавра, живший в раннем меловом периоде 130 - 125 млн лет назад.
История находки


{kind=link}
Concavenator corcovatus
Голотип MCCM-LH 6666 был обнаружен в 2003 году в провинции Куэнка, Испания. Датируется барремским ярусом. В то время на месте Испании находилась субтропическая область, богатую болотами, обеспечившими исключительную сохранность образцов. Благодаря этому скелет конкавенатора является почти полным, исключая несколько позвонков и мелких костей. Вероятно, тело оказалось в неглубоком бассейне на берегу озера с высоким содержанием карбоната кальция и там, быстро покрывшись бактериальными матами, полностью обызвествился. При этом остатки ещё около недели высыхали в другом месте и были занесены в водоём с паводком, который повредил тело. После этого их покрыл бактериальный мат, надёжно сохранив все остатки.
Большинство остатков из Лас-Хойас, где был обнаружен конкавенатор, очень малы, и голотип этого теропода является самым крупными из всех. Он содержит 94,1% процент остатков из которых 51,52% находятся в изначальном положении.
Сохранились также некоторые мягкие ткани - кожа, мышцы, а также содержимое желудка - 20 позвонков меньшего животного. На мышцы приходится 37,28% от всех уцелевших мягких тканей. Повреждения скелета включают отсутствие левой передней конечности, фрагментированную левую заднюю конечность и повреждённый инструментом затылок. Для сборки окаменелости потребовалось два года и ещё около пяти лет для описания крупнейшего испанского теропода.
Конкавенатор был описан в 2010 году. Его название переводится как "Горбатый охотник из Куэнки".
В 2018 году были подробно изучены череп и передние конечности. Эти исследования были сделаны в докторской диссертации Елены Куэста-Фидальго. Она также подробно изучила чешую задних конечностей с помощью ультрафиолетового излучения в 2015-16 годах.
Описание


{kind=link}
Конкавенатор был среднего размера, примерно 6 метров (20 футов) в длину.[1] В 2016 году Грегори Пол оценил конкавенатора в 5 метров в длину при весе в 300 фунтов. Куэста-Фидальго в 2018 году же оценила его вес в 442 кг.
Это был примитивный кархародонтозавр, обладающий несколькими уникальными особенностями. Два чрезвычайно высоких позвонка (одиннадцатый и двеннадцатый) впереди бедра формировали высокий, но узкий и острый гребень (возможно поддерживали горб) на спине динозавра. Роджер Бенсон предположил, что горб мог играть роль отличительного признака у самцов, а испанские учёные предположили, что он выполнял терморегуляционную функцию.
Отличительные черты конавенатора были изначально выявлены в 2010 году, но потом их посчитали недействительными. В первой публикации 2018 года были выявлены три уникальные особенности черепа: углубления на носовой кости соединены системой бороздок, задняя полость возле ноздрей удлинена и заострена, выступ над глазницей больше и внизу более округлый, чем у других кархародонтозавров. Последующая публикация указала на ещё четыре аутапоморфии: дельтопектральный гребень плечевой кости спереди имеет понижение, отросток локтевой кости короче, чем тело кости, третья фаланга третьего пальца равна по длине первой и второй вместе взятым, нижний отросток передней части подвздошной кости не до конца изогнут.
Череп


{kind=link}
Череп слегка повреждён, из-за чего нельзя выявить точное количество зубов. Достигает в длину около 50 см, профиль слегка удлинён и выпуклый, перед глазницей находится небольшой закруглённый выступ. Тяжёлые выросты, как у более поздних кархародонтозаврид, отсутствуют, как и слияние слёзной и предглазничной кости. Перед ноздрёй находится глубокая высокая ямка, глубоко входящая в премаксилу, постепенно сужаясь. Гребень над ямкой толстый, вертикальный, уходит вниз к анторбитальной фенестре. Сама фенестра большая, внизу ограничена чётким горизонтальным гребнем. На верхней челюсти четыре венозных канала не находились в горизонтальной бороздке. Восходящая ветвь верхней челюсти имеет выемку в верхней внешней части. Отверстие перед анторбитальной фенестрой большой, овальное и глубокое. Присутствует и маленькое плохо сохранившееся отверстие, что может быть промаксиллярной фенестрой. Носовые кости имеют неровные боковые края, под которыми расположены ряды из четырёх углублений, три из них соединены канавками. Слезная кость имеет L-образную форму. Передний край нисходящей ветви слезной кости имеет глубокую впадину между внутренней стенкой и внешней стенкой. Наружная стенка пересекает анторбитальную фенестру на середине длины слёзной кости. Верхняя часть загнута назад и несёт невысокий прямоугольный выступ. Под выступом и на нисходящей ветви расположены пневматические отверстия. На нисходящей ветви расположен небольшой выступ, проникающий в глазницу. Лобные кости плоские, сросшиеся с теменными сзади. На границе между префронтальной и заглазничной частью имеется небольшая бороздка. Слёзная кость отделяет префронтальную кость от заглазничной. Заглазничная кость имеет массивный грубый высокий прямоугольный выступ сразу за слезной костью. Выступ проходит высоко над глазницей сбоку, угловатый вверху и закруглённый внизу. Вверху виден костный шов, свидетельствующий о включении глазничных костей. Нисходящая ветвь заглазничной кости имеет на переднем крае довольно высокий округлый бугорок, придающий глазнице форму ключа. Глаз был выше выпуклости. В отличие от того, что было заявлено в 2010 году, передняя ветвь скулы проходит ниже анторбитального отверстия и отделяет верхнюю челюсть от слезной кости. Скула с прямым нижним краем и без выраженного горизонтального гребня.
Зубы типичные, кинжаловидные, уплощённые с боков. На эмали верхнечелюстных зубов есть морщинки, которые часто встречаются у многих теропод, но не так ярко выраженные, как у более поздних кархародонтозаврид. Зазубренные поверхности имеют по три острия в форме долота на зубце.
Нижние челюсти довольно плоские, а длинные зубы верхней челюсти выступают ниже линии нижней челюсти. У нижней челюсти спереди нет «подбородка». Также в нижней челюсти невозможно определить количество зубов. Дентарий снаружи гладкий. Боковой сустав подвижный, не жесткий.
Посткраниальный скелет
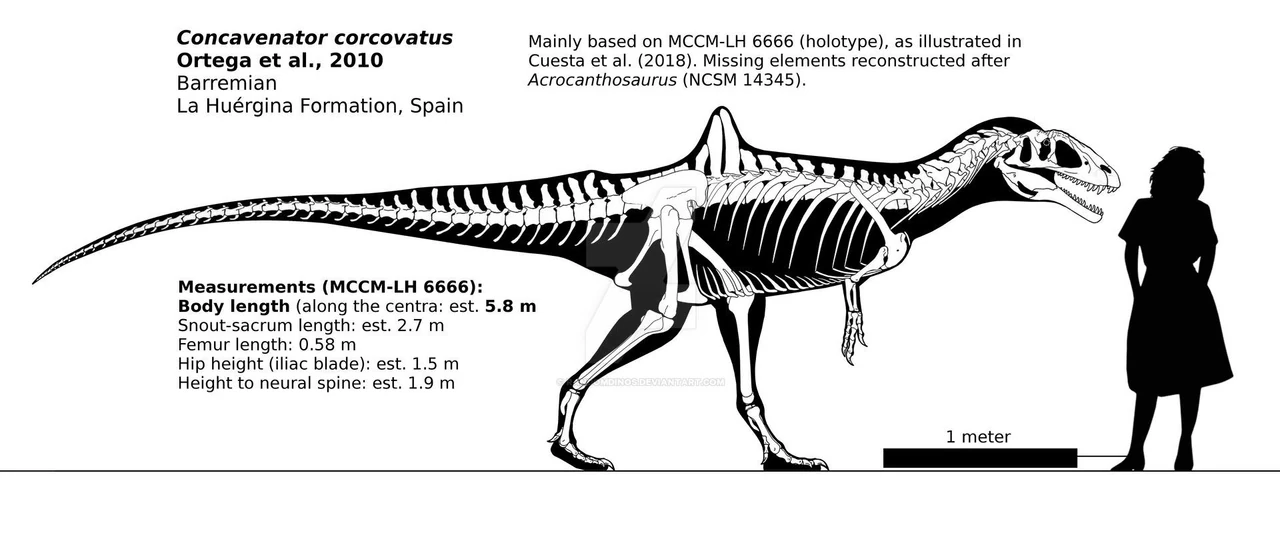

{kind=link}
Concavenator corcovatus от RandomDinos
Позвоночник состоял из 10 шейных позвонков, 13 позвонков, 5 крестцовых позвонков и не менее 39 хвостовых позвонков, хотя в общей сложности их число должно было превышать 40. Из-за того, что окаменелость расплющена, невозможно определить, насколько пневматизированными были позвонки.
Атлант конкавенатора имеет длинный и узкий эпипофиз, выступающий у основания заднего суставного отростка, отличный от короткого эпипофиза аллозавра и широкого эпипофиза заурофаганакса. Второй шейный позвонок имеет уплощённый в поперечнике остистый отросток, что отличает его от мапузавра и гиганотозавра.
Самой удивительной особенностью конкавенатора является небольшой парус на спине. Единственный ящер с подобным строением - альтиспинакс, у всех остальных теропод с парусами он проходил вдоль всей спины. У конкавенатора только 11 и 12 спинные позвонки имеют удлинённые невральные шипы, выступающие примерно на 40 см. Десятый позвонок имеет несколько удлиненный выступ, профиль которого, однако, приподнят кзади, так что постепенный переход все же в определенной степени образуется. Однако на шести крестцовых позвонках таза, тринадцатом позвонке и далее остистые отростки очень низкие. Это приводит к узкому, но высокому парусу прямо перед бедром. На передних хвостовых позвонках шипы снова начинают удлиняться, в результате чего образуется нижний вторичный хвостовой гребень. Функции паруса неизвестны: были предположения, что он играл роль в терморегуляции, помогал накапливать жир, но самую вероятную гипотезу предположил Пол Серено. Он посчитал, что парус играл роль в социальном поведении конкавенаторов, помогая привлечь внимание самки или отогнать потенциального соперника. Однако пока мы знаем только один образец конкавенатора, так что утверждать что-либо сложно. Разве что можно быть уверенным, что парус не участвовал в терморегуляции:в кости нет следов расширенной системы кровеносных сосудов. Кроме того, уникальное производное свойство или аутапоморфия конкавенатора заключается в том, что одиннадцатый и двенадцатый позвонки имеют дополнительные гребни поверх центра основания боковых шипов.
Оперение


{kind=link}
У конковенатора на локтевых костях были обнаружены небольшие шишки, сходные с присоединительными местами перьев, которые характерны для птиц и других пернатых тероподов, таких как велоцираптор, и птицетазовых динозавров, вроде тяньюйлуна и пситтакозавра. Данные шишки образовывали связки для прикрепления фолликул пера. Поскольку чешуя не может образовывать фолликулы, идея о длинных иглоподобных чешуях на передних конечностях сразу отпала. Вместо этого была предложена гипотеза о наличии примитивных, похожих на перья, структур.
До обнаружения конкавенатора ни у одного представителя аллозавроид следов перьевых структур ранее не находили, в то время как у параллельной ветви тероподов — целурозавров — они имеются. Если бугорки птицетазовых динозавров не связаны с перьями, наличие этих структур у конкавенатора показало бы, что перья начали появляться на более ранних этапах эволюции и у более примитивных представителей клады неотетанур, что отодвигает предложительный момент возникновения перьев в середину юрского периода, к отметке возраста в 175—161 млн лет. Тем не менее, на руках не удалось обнаружить следов каких-либо покровов, хотя остальные участки тела сохранили отпечатки чешуи, включая широкие прямоугольные чешуйки на нижней стороне хвоста, птичьи чешуйки на ногах и подошвенные чешуи на нижней стороне ног.


Некоторые эксперты со скептицизмом отнеслись к птицеподобной природе шишек на конечностях. Даррен Нейш из блога Tetrapod Zoology указал, что бугорки располагались слишком далеко и разрозненно для перьевых шишек. Он также отметил, что многие животные имеют подобные структуры вдоль межмышечных линий, которые служат для крепления сухожилий. Эту версию поддержал Кристиан Форт в 2014 году. Гипотеза о том, что бугорки на локтевой кости нужны были для крепления мышц, была подробно исследована, а результаты представлены на заседании Общества палеонтологии позвоночных в 2015 году. Елена Куэста-Фидальго вместе с авторами описания конкавенатора попытались реконструировать его мускулатуру предплечья, чтобы определить, могли ли шишки служить в качестве межмышечных гребней. Когда они определили место расположения мышц, оказалось, что бугорки не могли располагаться между ними. Выдвинутая в процессе исследования гипотеза о креплении локтевой мышцы к бугоркам оказалась маловероятной, так как обычно эта мышца крепится к гладкой костной поверхности без каких-либо специальных мест крепления. Авторы сошлись во мнении, что первоначальная интерпретация шишек как мест для крепления перьев наиболее вероятна. Также они отметили, что, несмотря на необычное место расположения бугорков, такая же структура иногда встречается и у современных птиц, вроде камышниц.


{kind=link}
Реконструкция конкавенатора с парусом и чешуёй на руках
Тем не менее, некоторые палеонтологи не согласились с выводами Куэста-Фидальго, Ортеги и Санса. Микки Мортимер указал, что локтевые бугорки, по-видимому, на самом деле находятся на переднебоковой поверхности кости, а не на заднебоковой, как посчитала группа Фидальго. Он также высказал мнение, что крокодилы подходят лучше для реконструкции мышц конкавенатора и других неманирапторовых тероподов, чем птицы. Мортимер утверждал, что локтевые бугорки, скорее всего, являются частью межмышечной линии. Андреа Кау сравнил предплечья конкавенатора, аллозавра и акрокантозавра, выяснив, что у голотипа первого оно оказалось вывихнутым и что на самом деле бугорки находились на передней стороне конечности. Из-за того, что бугорки не находоились на одной стороне локтевой кости, Кау утверждал, что они не могут быть гомологичны перьевым шишкам манирапторов. Кроме того, против этой гипотезы говорили и другие факты - бугорки конкавенатора расположены неравномерно и соединены вогнутым гребнем, в то время как у птиц они расположены равномерно и по отдельности. Кау также добавил, что филогенетически маловероятно наличие перьев у аллозавроидов, поскольку он считал их уникальной чертой манирапторов. Интерпретация Кау заключалась в том, что эти бугорки служили для крепления мышц, указывая на аналогичную структуру у балаура.


{kind=link}
В 2018 году Куэста-Фидальго опубликовала свою докторскую работу, посвящённую анатомии конкавенатора, которая утверждала, что локтевая кость сохранилась в боковом положении, что указывало на расположение бугорков на задней стороне руки, а не передней. Также было отмечено, что проксимальный конец локтевой кости был повреждённым, с многочисленными переломами и от части истёртым. Это привело к тому, что некоторые части кости были смещены. Например, у аллозавра и акрокантозавра латеральный отросток находился на передней части, а у конкавенатора оказался смещён назад. Поэтому при жизни костные бугорки были направлены вперёд и вбок. Тем не менее, весомых доказательств для предположения о перемещении кости в переднее положение. Кроме того, из-за повреждений и искажений пропорций конечности конкавенатора не получится точно сравнить с конечностями других аллозавроидов. Если кость сохранилась повёрнутой на бок, то бугорки не могли поддерживать мышцы. Кроме того, Мэтт Мартынюк отметил, что наличие перьев ещё у примитивных тероподов вполне вероятно. В качестве аргумента он использовал бескрылых дилунов и компосгнатид, у которых были обнаружены полноценные перья, не участвующие в создании широких, похожих на крылья структур. Однако Мартынюк отметил, что отсутствие у дилунов и компсогнатид крыльев, возможно, было независимой реверсией, коррелирующей с уменьшением передних конечностей в этих линиях, аналогично случаю со многими современными нелетающими птицами.
Систематика
Кладистические анализы показали разное положение конкавенатора: у истоков кархародонтозавров, являясь родственником кархародонтозаврид и неовенаторид, и в качестве базального члена кархародонтозаврид.
Анализ 2013 года:
| |||||||||||||||||||||||||||||||||||||||||||||||||
Исследования 2018 года подтвердили базальное место конкавенатора среди кархародонтозаврид. У него отсутствуют признаки прогрессивных кархародонтозаврид: орнамент на внешней стороне верхней челюсти, выступ на нижней стороне скулы, соединение предглазничной и слёзной кости, "подбородок" на нижней челюсти, уменьшенный расширяющийся выступ нижней челюсти и удлинённая квадратная точка зубного ряда.
Анализ, сделанный при описании кархародонтозаврида мераксеса:
| Carcharodontosauridae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Палеоэкология
Конкавенатор обитал в субтропической местности с большим количеством болот. Средняя температура колебалась от 20°C зимой до 44°C летом. Большинство местных останков принадлежат болотным животным, поэтому считается, что конкавенатор попал в болото случайно уже после смерти.
Источники
Галерея
")
")
")
")
")
")
")
")
")
")
.jpg "Maxresdefault (22).jpg (57 Кб)")
| Аветероподы | ||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
Газозавр(?) • Ловринанозавр(?)
|