![]()
Чаохузавр (лат. Chaohusaurus) — ранний представитель ихтиозавроморф, обитал на территории Китая в триасовом периоде, около 251—247 млн лет назад.
История находки
Ископаемые окаменелости были обнаружены при строительстве железной дороги в провинции Аньхой, на востоке Китая. В 1972 году китайские ученые Ян Чжунцзян и Донг Чжиминг описали эту находку под видом Chaohusaurus geishanensis. Голотип IVPP V 4001 состоит из частичного скелета, содержащего череп и передний торс. В 1985 году Чэн Лизху назвал два дополнительных вида на основе окаменелостей, обнаруженных в той же геологической формации: Anhuisaurus chaoxianensis и Anhuisaurus faciles, однако позже выяснилось, что название рода уже используется в классификации ящериц. В 1991 году Жан-Мишель Мазин переименовал этот род в Chensaurus, по имени Чена, сделавшего первичное описание. В 1998 году исследование Рёсуке Мотани выявило, что эти окаменелости представляют собой останки несовершеннолетних особей, представляющих собой различные стадии роста Chaohusaurus.

{kind=link}
Chaohusaurus geishanensis (AGM CH-628-22)
В 2013 году был назван второй вид Chaohusaurus zhangjiawanensis, название вида относится к месту его обнаружения вблизи деревни Чжанцзявань. В 2014 году сообщалось о трех дополнительных образцах: AGM I-1, AGM CHS-5 и AGM CH-628-22, первый из которых содержал останки трех эмбрионов. Новые находки являются частью недавних открытий китайско-итальянского палеонтологического проекта.

{kind=link}
Сравнение размеров с человеком.
Описание
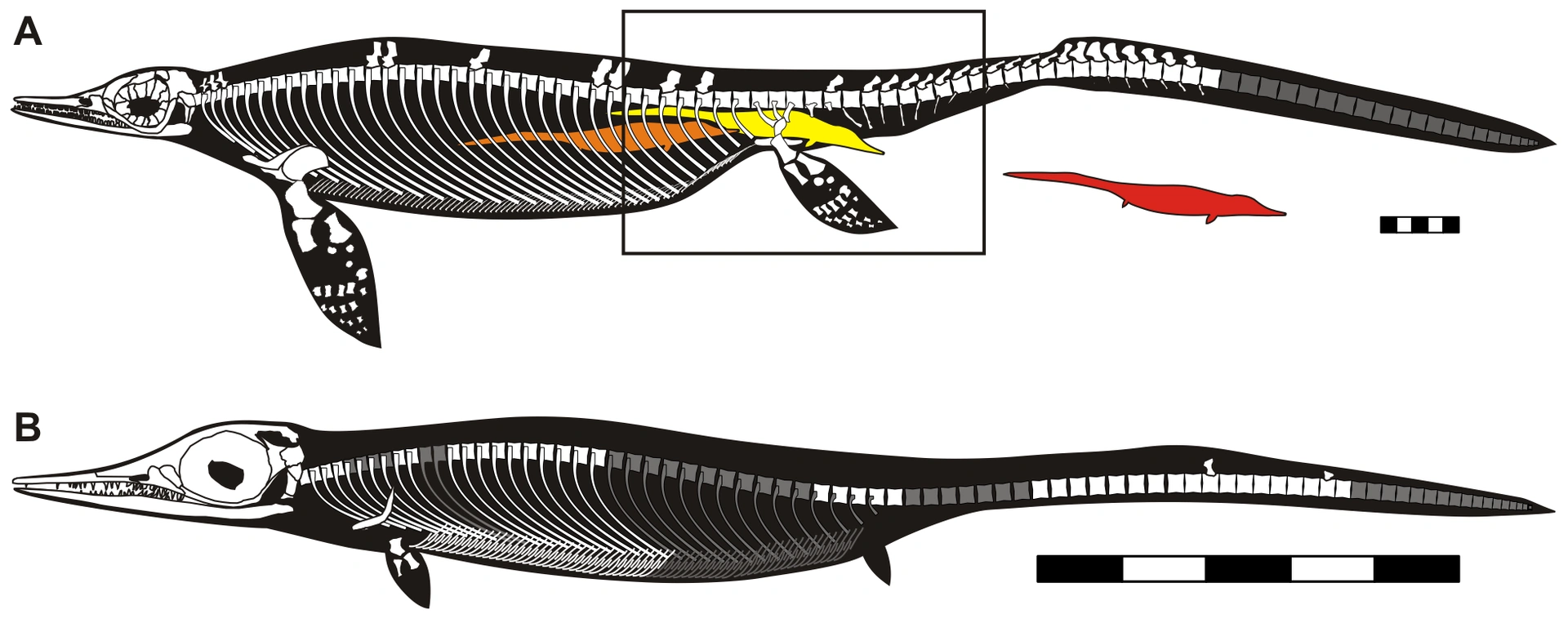

{kind=link}
Реконструкция взрослой (А) и ювенильной особи (В)
Заметная базальная черта — короткая голова; в более поздних формах он будет длиннее ствола. Кроме того, морда относительно короткая, всего примерно в два раза длиннее части за глазницей. Носовые не выходят назад, чем глазницы. Шов между носовой костью и лобной костью ориентирован поперечно. С префронтальной части к верхнему переднему краю глазницы нависает фланец, возможно, для защиты глаз. Лобная часть является частью верхнего края глазницы и, следовательно, лишена бокового упора. Лобная кость примерно такого же размера, как и теменная. На виде сверху задний край крыши черепа с выемкой. Теменный глаз расположен между теменными костью, а не между лобными костью, как у более поздних форм. Чешуйчатая кость — крупный элемент, размером с квадратично-скуловую кость, с которой он прочно срастается. Базисфеноид узкий. Культовидный отросток парасфеноида сзади постепенно сливается с основным телом, а не через узкую талию, как у более поздних форм. Нёбо не было прочно прикреплено к базиптеригоиду, что позволяло рылу некоторое вертикальное движение относительно остальной части черепа. У большинства ихтиозавров такое движение было невозможно. Отверстие между крыловидными мышцами узкое, щелевидное, не широкое. Крыловидные кости не закрывают заднюю нижнюю часть черепной коробки. Шея относительно длинная. Хвост удлиненный, примерно такой же длины, как голова, шея и туловище вместе взятые. Позвонки основания хвоста также удлинены, их длина примерно равна их высоте. Самые длинные хвостовые шипы расположены скорее впереди, что указывает на то, что возможный хвостовой плавник должен быть более горизонтальным, чем у более поздних форм. Ключицы имеют широкий внутренний выступ. Лопатка короткая, ее ширина больше длины. Голова по-прежнему отчетливая, но спинного вертела еще нет. На нижнем конце плечевая кость имеет большую фасетку, которая контактирует с лучевой костью, чем с локтевой. Верхний конец локтевой кости уже нижнего, не равен ему по размеру. В целом кости предплечья, в том числе и кисти, довольно удлиненные, не переходят в диски. Локтевая и лучевая кость все еще имеют стержень, как у наземных животных. На запястье гороховидная форма размером примерно с локтевую кость запястья ненамного меньше или отсутствует. Пятая пястная кость имеет выпуклый задний край и длиннее четвертой дистальной части запястья. Осталось пять пальцев, без уменьшения первого пальца. Таким образом, отсутствует полидактилия, лишнее количество пальцев. Гиперфаланги, лишние фаланги также отсутствуют. Формула фаланги 2-3-4-4-2. Верхние фаланги относительно удлиненные, их длина больше ширины. Нижние фаланги все еще имеют окостенение ниже надхрящницы хряща и имеют выемки по краям. Лобковая кость перфорирована затылочным отверстием, закрытым при виде сбоку и расположенным на некотором расстоянии от заднего края тела. Задний плавник примерно такого же размера, как передний, но не меньше. В бедренной кости фасетка, контактирующая с большеберцовой костью, такая же большая и достигает вниз, как фасетка, контактирующая с малоберцовой костью. Между большеберцовой и малоберцовой костями все еще есть пространство, и обе кости, хотя и уплощенные, относительно удлинены с четким стержнем. То же самое и с плюсневыми костей, которые имеют форму цилиндра. Осталось еще пять пальцев на ногах, опять же довольно удлиненных и в форме песочных часов.
Ранняя производная черта — относительно короткие шипы на хвосте. Плечевая кость имеет выступ на переднем крае, но вторично не сокращается. На фланце есть небольшая выемка. Пятая плюсневая кость укорачивается, а первая еще короче. Пятый палец короче первого пальца.
У Chaohusaurus ширина базисфеноида составляет около 63 % его длины. У Chaohusaurus есть сочетание более острых и выпуклых зубов, которые, вероятно, развились отдельно.
Второй вид, Chaohusaurus zhangjiawanensis, обладает некоторыми отличительными чертами. Череп довольно плоский. У туловищных позвонков хорошо развиты поперечные отростки. Префронтальная кость соприкасается с постфронтальной костью, исключая фронтальную кость от края глазницы, что является производным признаком. Пяточная кость больше. Первое крестцовое ребро имеет расширенный внешний конец. Второе крестцовое ребро имеет сужающийся конец.
Филогения
Chaohusaurus был в первоначальном описании 1972 года отнесен к Omphalosauridae. Современные исследователи либо воздерживаются от включения в семейство, либо относят к Grippiidae. Chaohusaurus — возможный сестринский вид Grippia в Grippidia.
Точное размещение Chaohusaurus зависит от выбора определений. Согласно филогенезу Мотани, это базальная ихтиоптеригия, но не ихтиозавр:
| Ихтиоптеригии |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Однако, согласно Maisch & Matzke, чаохузавр является базальным ихтиозавром:
| Ichthyosauria |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
Кладистический анализ, опубликованный с описанием Chaohusaurus zhangjiawanensis, не обнаружил, что этот таксон является сестринским видом Chaohusaurus geishanensis, более близким к Grippia. Несмотря на то, что это сделало бы род парафилетическим, C. zhangjiawanensis тем не менее был помещен в Chaohusaurus из-за морфологического сходства с типовым видом.
Реконструкция


{kind=link}
AGM I-1
Один экземпляр из числа обнаруженных в 2014 году, AGM I-1, также содержал останки двух эмбрионов и одного новорожденного. Таким образом, Chaohusaurus, подобно более поздним ихтиозаврам, рождает своих детенышей в воде живородящим образом. Однако по ориентации как эмбрионов внутри тела матери, так и новорожденного, было ясно, что детеныши выходили из родового канала головой вперед. Это отличается от метода, используемого большинством современных морских живородящих амниот, которые рождают детёнышей хвостом вперед, чтобы они не задохнулись. Многие более молодые экземпляры ихтиозавров ранее были обнаружены с эмбрионами в обоих положениях, поэтому не удалось решить, какое из них является нормальным. Motani et alii (2014) пришли к выводу, что, поскольку Chaohusaurus — очень базальная форма, это дает убедительные доказательства того, что, по крайней мере первоначально, ихтиоптеригийские детеныши рождались с головой вперед. Позднее этот ранний метод мог быть изменен, поскольку он приводил к слишком высокой смертности. Они также назвали это доказательством наличия живорождения у наземных предков ихтиозавров. Таким образом, AGM I-1 в 2014 году представлял собой самую старую из известных живородящих рептилий.
Источники
Литература
- Mazin, J.-M.; Suteethorn, V.; Buffetaut, E.; Jaeger, J.-J.; Helmckeingavat, R. (1991). «Preliminary description of Thaisaurus chonglakmanii n. g., n. sp., a new ichthyopterygian (Reptilia) from the Early Triassic of Thailand». Comptes Rendus de l’Académie des Sciences, Série II. 313: 1207—1212.
- Liezhu, Chen (1985). «[Ichthyosaurs from the lower Triassic of Chao County]». Anhui Regional Geology of China. 15: 139—146.